Viskoelasticitet är en egenskap hos mänskligt blod som främst beror på den elastiska energi som lagras i de röda blodkropparnas deformation när hjärtat pumpar blodet genom kroppen. Den energi som hjärtat överför till blodet lagras delvis i den elastiska strukturen, en annan del avdispenseras av viskositeten och den återstående energin lagras i blodets kinetiska rörelse. När hjärtats pulsation beaktas blir en elastisk regim tydligt tydlig. Det har visats att det tidigare begreppet blod som en rent viskös vätska var otillräckligt eftersom blod inte är en vanlig vätska. Blod kan mer korrekt beskrivas som en fluidiserad suspension av elastiska celler (eller en sol).
De röda blodkropparna upptar ungefär hälften av blodets volym och har elastiska egenskaper. Denna elastiska egenskap är den största bidragande faktorn till blodets viskoelastiska beteende. Den stora volymprocenten av röda blodkroppar vid en normal hematokritnivå lämnar lite utrymme för cellrörelse och deformation utan att interagera med en angränsande cell. Beräkningar har visat att den maximala volymprocenten för röda blodkroppar utan deformation är 58 %, vilket ligger inom intervallet för normalt förekommande nivåer. På grund av det begränsade utrymmet mellan de röda blodkropparna är det uppenbart att för att blodet ska kunna flöda spelar betydande interaktion mellan cellerna en nyckelroll. Denna växelverkan och tendensen till att cellerna aggregerar är en viktig faktor som bidrar till blodets viskoelastiska beteende. Deformation och aggregering av röda blodkroppar är också kopplad till flödesinducerade förändringar i arrangemanget och orienteringen som en tredje viktig faktor för dess viskoelastiska beteende. Andra faktorer som bidrar till blodets viskositet är plasmaviskositeten, plasmasammansättningen, temperaturen och flödeshastigheten eller skjuvningshastigheten. Tillsammans gör dessa faktorer människoblodet viskoelastiskt, icke newtonskt och thixotropt.
När de röda blodkropparna är i vila eller vid mycket små skjuvningshastigheter tenderar de att aggregera och stapla ihop sig på ett energimässigt gynnsamt sätt. Attraktionen tillskrivs laddade grupper på cellernas yta och förekomsten av fibrinogen och globuliner. Denna aggregerade konfiguration är ett arrangemang av celler med minsta möjliga deformation. Vid mycket låga skjuvningshastigheter domineras blodets viskoelastiska egenskaper av aggregeringen och cellernas deformbarhet är relativt obetydlig. När skjuvningshastigheten ökar börjar aggregatens storlek minska. Vid ytterligare ökning av skjuvningshastigheten kommer cellerna att omorganiseras och orientera sig för att skapa kanaler för plasman att passera igenom och för cellerna att glida. I detta intervall med låg till medelhög skjuvningshastighet vickar cellerna i förhållande till granncellerna och möjliggör flödet. Aggregationsegenskapernas inflytande på viskoelasticiteten minskar och de röda blodkropparnas deformbarhet börjar öka. När skjuvningshastigheten blir stor kommer de röda blodkropparna att sträcka sig eller deformeras och anpassa sig till flödet. Cellskikt bildas, separerade av plasma, och flödet tillskrivs nu lager av celler som glider på lager av plasma. Cellskiktet gör det lättare för blodet att flöda och därmed minskar viskositeten och elasticiteten. Blodets viskoelasticitet domineras av de röda blodkropparnas deformbarhet.
MaxwellmodellenRedigera
Maxwellmodellen gäller Maxwellvätskor eller Maxwellmaterial. Materialet i Maxwellmodellen är en vätska, vilket innebär att det respekterar kontinuitetsegenskaperna för konservativa ekvationer : Fluider är en delmängd av materiens faser och omfattar vätskor, gaser, plasmor och i viss mån plastiska fasta ämnen. Maxwellmodellen är gjord för att uppskatta lokala konservativa värden för viskoelasticitet genom ett globalt mått i modellens integralvolym som kan överföras till olika flödessituationer. Blod är ett komplext material där olika celler, t.ex. röda blodkroppar, är diskontinuerliga i plasma. Deras storlek och form är också oregelbunden eftersom de inte är perfekta sfärer. De röda blodkropparna är inte identiskt fördelade i en blodprovsvolym, eftersom de vandrar med hastighetsgradienter i riktning mot de områden där hastigheten är högst, vilket är den berömda Fåhræus-Lindqvist-effekten, och aggregerar eller separerar sig i skida- eller pluggflöden som beskrivs av Thurston. Typiskt sett betraktar Maxwellmodellen som beskrivs nedan materialet (enhetlig blå färg) som en perfekt distribuerad partiklarvätska överallt i volymen (i blått), men Thurston avslöjar att paket av röda blodkroppar, pluggar, är mer närvarande i höghastighetsområdet, om y är höjderiktningen i Maxwellmodellens figur, (y~H) och det finns ett lager med fria celler i området med lägre hastighet (y~0), vilket innebär att den plasmabaserade vätskefasen som deformeras enligt Maxwellmodellen är ansträngd till följd av inre foder som helt och hållet undgår Maxwells analytiska modell.
I teorin beter sig en vätska i en Maxwellmodell exakt likadant i vilken annan flödesgeometri som helst, t.ex. rör, roterande celler eller i viloläge. Men i praktiken varierar blodets egenskaper med geometrin och blodet har visat sig vara ett otillräckligt material för att studeras som en vätska i vanlig mening. Maxwellmodellen ger alltså tendenser som måste kompletteras i verkliga situationer, följt av Thurstonmodellen i ett kärl när det gäller fördelningen av celler i skida och pluggflöden.
Om man betraktar en liten kubisk blodvolym med krafter från hjärtats pumpning och skjuvkrafter från gränserna. Förändringen av kubens form kommer att ha 2 komponenter:
- Elastisk deformation som kan återställas och lagras i blodets struktur.
- Slipning som är förknippad med en kontinuerlig tillförsel av viskös energi.
När kraften avlägsnas skulle kuben delvis återställas. Den elastiska deformationen är omvänd men inte glidningen. Detta förklarar varför den elastiska delen endast är märkbar i instabila flöden. I stabilt flöde kommer glidningen att fortsätta att öka och mätningarna av icke tidsvarierande kraft kommer att försumma bidragen från elasticiteten.

Figur 1 kan användas för att beräkna följande parametrar som är nödvändiga för utvärderingen av blodet när en kraft utövas.
Skjuvspänning: τ = F A {\displaystyle \tau ={\frac {F}{A}}}

Skjuvspänning: γ = D H {\displaystyle \gamma ={\frac {D}{H}}}}

Skjuvhastighet: γ ˙ = V H {\displaystyle {\dot {\gamma }}={\frac {V}{H}}}}

Ett sinusformigt tidsvarierande flöde används för att simulera pulsationen i ett hjärta. Ett viskoelastiskt material som utsätts för ett tidsvarierande flöde kommer att resultera i en fasvariation mellan τ {\displaystyle \tau }

och γ {\displaystyle \gamma }

som representeras av ϕ {\displaystyle \phi }

. Om ϕ = 0 {\displaystyle \phi =0}

är materialet rent elastiskt eftersom spänningen och töjningen är i fas, så att den ena reaktionen orsakad av den andra är omedelbar. Om ϕ {\displaystyle \phi }
= 90° är materialet ett rent visköst eftersom töjningen släpar efter spänningen med 90 grader. Ett viskoelastiskt material kommer att ligga någonstans mellan 0 och 90 grader.
Den sinusformade tidsvariationen är proportionell mot e i ω t {\displaystyle e^{i\omega t}}

. Därför beskrivs storleks- och fasförhållandet mellan spänning, töjning och skjuvningshastighet med hjälp av detta förhållande och en radiär frekvens, ω = 2 π f {\displaystyle \omega =2\pi f}

var f {\displaystyle f}

är frekvensen i hertz. Skjuvspänning: τ ∗ = τ e – i ϕ {\displaystyle \tau ^{*}=\tau e^{-i\phi }}

Skjuvspänning: γ ∗ = γ e – i π 2 {\displaystyle \gamma ^{*}=\gamma e^{-i{\frac {\pi }{2}}}}

Skjuvhastighet: γ ˙ ∗ = γ ˙ e – i 0 {\displaystyle {\dot {\gamma }}^{*}={\dot {\gamma }}e^{-i0}}

Komponenterna av den komplexa skjuvspänningen kan skrivas som:
τ ∗ = τ ′ – i τ ″ {\displaystyle \tau ^{*}=\tau ’-i\tau ”}

Varvid τ ′ {\displaystyle \tau ’}

är den viskösa spänningen och τ ″ {\displaystyle \tau ”}

är den elastiska spänningen.Den komplexa viskositetskoefficienten η ∗ {\displaystyle \eta ^{*}}

kan hittas genom att ta förhållandet mellan den komplexa skjuvspänningen och den komplexa skjuvhastigheten: η ∗ = τ ∗ γ ˙ ∗ = ( τ ′ γ ˙ + i τ ″ γ ˙ ) = η ′ + i η ″ {\displaystyle \eta ^{*}={\frac {\tau ^{*}}}{{\dot {\gamma }}^{*}}}}=({\frac {\tau ’}{\dot {\gamma }}}+i{\frac {\tau ”}{\dot {\gamma }}})=\eta ’+i\eta ”}

På samma sätt kan den komplexa dynamiska modulen G erhållas genom att ta förhållandet mellan den komplexa skjuvspänningen och den komplexa skjuvdeformationen.
G = τ ∗ γ ∗ = ( τ ″ γ + i τ ′ γ ) {\displaystyle G={\frac {\tau ^{*}}}{\gamma ^{*}}}=({\frac {\tau ”}{\gamma }}+i{\frac {\tau ’}{\gamma }})}

Om man relaterar ekvationerna till vanliga viskoelastiska termer får man lagringsmodulen G’ och förlustmodulen G”.
G = G ′ + i G ″ {\displaystyle G=G’+iG”}

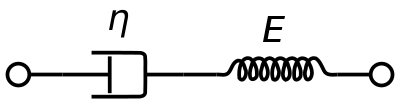
En viskoelastisk Maxwell-materialmodell används ofta för att representera blodets viskoelastiska egenskaper. Den använder rent viskösa dämpare och en rent elastisk fjäder som är kopplade i serie. Analysen av denna modell ger den komplexa viskositeten i termer av dashpotkonstanten och fjäderkonstanten.
η ∗ = η d a s h 1 + i ω ( η d a s h E s p r i n g ) = η ′ – i η ″ {\displaystyle \eta ^{*}={\frac {\eta _{dash}}{1+i\omega ({\frac {\eta _{dash}}{E_{fjäder}}})}}=\eta ’-i\eta ”}

Oldroyd-B modellEdit
En av de mest använda konstitutiva modellerna för blodets viskoelasticitet är Oldroyd-B modellen. Det finns flera varianter av Oldroyd-B:s icke-newtonska modell som karakteriserar skjuvförtunningsbeteendet på grund av aggregering och dispersion av röda blodkroppar vid låg skjuvningshastighet. Här betraktar vi en tredimensionell Oldroyd-B-modell kopplad till impulsekvationen och den totala spänningstensorn. Ett icke newtonskt flöde används, vilket garanterar att blodets viskositet μ ( h , d ) {\displaystyle \mu (h,d)}

är en funktion av kärldiametern d och hematokriten h. I Oldroyd-B-modellen ges förhållandet mellan skjuvspänningensorn B och orienteringsspänningensorn A av följande:
S + γ = μ ( h , d ) – g A + C 1 ( g A – C 2 I μ ( h , d ) 2 ) {\displaystyle S+\gamma \left=\mu (h,d)\left-gA+C_{1}\left(gA-{\frac {C_{2}I}{\mu (h,d)^{2}}}}\right)}

där D/Dt är materialderivatan, V är vätskans hastighet, C1, C2, g, γ {\displaystyle \gamma }
är konstanter. S och B definieras på följande sätt: S = μ B + g A {\displaystyle S=\mu B+gA}

B = Δ V + ( Δ V ) T {\displaystyle B=\Delta V+(\Delta V)^{T}}

Viskoelasticitet hos röda blodkropparEdit
Röda blodkroppar utsätts för intensiv mekanisk stimulering från både blodflödet och kärlväggarna, och deras reologiska egenskaper är viktiga för deras effektivitet när det gäller att utföra sina biologiska funktioner i mikrocirkulationen. Röda blodkroppar i sig själva har visat sig uppvisa viskoelastiska egenskaper. Det finns flera metoder som används för att undersöka de mekaniska egenskaperna hos röda blodkroppar, t.ex:
- mikropipettaspiration
- mikrointryck
- optisk pincett
- högfrekventa elektriska deformationstester
Dessa metoder arbetade för att karaktärisera de röda blodkropparnas deformationsförmåga i termer av skjuv-, böjnings-, areaexpansionsmoduler och relaxationstider. De kunde dock inte utforska de viskoelastiska egenskaperna. Andra tekniker har genomförts, t.ex. fotoakustiska mätningar. Denna teknik använder en laserstråle med en enda puls för att generera en fotoakustisk signal i vävnader och signalens avklingningstid mäts. Enligt teorin om linjär viskoelasticitet är avklingningstiden lika med viskositets-elasticitetskvoten och därför kunde man få fram viskoelasticitetsegenskaperna hos de röda blodkropparna.
En annan experimentell teknik som användes för att utvärdera viskoelasticiteten bestod i att använda ferromagnetiska pärlor som var bundna till en cellyta. Krafter appliceras sedan på den magnetiska pärlan med hjälp av optisk magnetisk vridcytometri, vilket gjorde det möjligt för forskarna att utforska de röda blodkropparnas tidsberoende reaktioner.
T s ( t ) {\displaystyle T_{s}(t)}

är det mekaniska vridmomentet per enhet pärlvolym (spänningsenheter) och ges av: T s ( t ) = c H cos θ {\displaystyle T_{s}(t)=cH\cos \theta }

där H är det applicerade magnetiska vridfältet, θ {\displaystyle {\theta }}

är vinkeln på pärlans magnetiska moment i förhållande till den ursprungliga magnetiseringsriktningen, och c är pärlkonstanten som hittas genom experiment som utförs genom att placera pärlan i en vätska med känd viskositet och applicera ett vridningsfält.
Den komplexa dynamiska modulen G kan användas för att representera förhållandet mellan oscillerande spänning och töjning:
G = G ′ + i G ″ {\displaystyle G=G’+iG”}
där G ′ {\displaystyle G’}

är lagringsmodulen och G ″ {\displaystyle G”}

är förlustmodulen: G ′ = σ 0 ε 0 ε 0 cos ϕ {\displaystyle G’={\frac {\sigma _{0}}{\varepsilon _{0}}}}\cos \phi }

G ″ = σ 0 ε 0 ε 0 sin ϕ {\displaystyle G”={\frac {\sigma _{0}}}{\varepsilon _{0}}}\sin \phi }

varvid σ 0 {\displaystyle \sigma _{0}}

och ε 0 {\displaystyle \varepsilon _{0}}

är amplituderna för spänning och töjning och ϕ {\\displaystyle \phi }
är fasförskjutningen mellan dem.

Utifrån ovanstående relationer bestäms komponenterna i den komplexa modulen från en slinga som skapas genom att jämföra förändringen av vridmomentet med förändringen av tiden, som bildar en slinga när den representeras grafiskt. Gränserna för T s ( t ) {\displaystyle T_{s}(t)}
– d(t) slingan och området A som avgränsas av T s ( t ) {\displaystyle T_{s}(t)}
– d(t)-slingan, som representerar energiförlusterna per cykel, används i beräkningarna. Fasvinkeln ϕ {\displaystyle \phi }
, lagringsmodul G’ och förlustmodul G blir då: ϕ = sin – 1 4 A π Δ T s Δ d {\displaystyle \phi =\sin ^{-1}{\frac {4A}{\pi \Delta T_{s}\Delta d}}}}

G ′ = Δ T s Δ d cos ϕ {\displaystyle G’={\frac {\Delta T_{s}}}{\Delta d}}}\cos \phi }

G ″ = Δ T s Δ d sin ϕ = 4 A π ω Δ d 2 {\displaystyle G”={\frac {\Delta T_{s}}}{\Delta d}}\sin \phi ={\frac {4A}{\pi \omega \Delta d^{2}}}}

där d är förskjutningen.
Hysteresen som visas i figur 3 representerar den viskoelasticitet som finns i röda blodkroppar. Det är oklart om detta är relaterat till molekylära fluktuationer i membranen eller metabolisk aktivitet som kontrolleras av intracellulära koncentrationer av ATP. Ytterligare forskning behövs för att fullt ut utforska denna interaktion och för att belysa de röda blodkropparnas underliggande viskoelastiska deformationsegenskaper.
Effekter av blodkärlRedigera
När man tittar på blodets viskoelastiska beteende in vivo är det nödvändigt att även beakta effekterna av artärer, kapillärer och vener. Blodets viskositet har ett primärt inflytande på flödet i de större artärerna, medan elasticiteten, som ligger i de röda blodkropparnas elastiska deformbarhet, har ett primärt inflytande i arteriolerna och kapillärerna. Det är viktigt att förstå vågutbredningen i artärväggarna, den lokala hemodynamiken och skjuvspänningsgradienten i väggen för att förstå mekanismerna för den kardiovaskulära funktionen. Arterieväggarna är anisotropa och heterogena och består av lager med olika biomekaniska egenskaper, vilket gör det mycket svårt att förstå de mekaniska influenser som artärerna bidrar med till blodflödet.
Medicinska skäl för en bättre förståelseRedigera
Från en medicinsk synvinkel blir betydelsen av att studera blodets viskoelastiska egenskaper uppenbar. Med utvecklingen av kardiovaskulära proteser såsom hjärtklaffar och blodpumpar krävs förståelse för pulserande blodflöde i komplexa geometrier. Några specifika exempel är effekterna av blodets viskoelastiska egenskaper och dess konsekvenser för testning av pulserande blodpumpar. Starka korrelationer mellan blodets viskoelasticitet och regionalt och globalt cerebralt blodflöde under kardiopulmonell bypass har dokumenterats.
Detta har också banat väg för att utveckla en blodanalog för att studera och testa proteser. Den klassiska analogin av glycerin och vatten ger en bra representation av viskositet och tröghetseffekter men saknar de elastiska egenskaperna hos verkligt blod. En sådan blodanalog är en vattenlösning av xantangummi och glycerin som utvecklats för att matcha både de viskösa och elastiska komponenterna i blodets komplexa viskositet.
Normala röda blodkroppar är deformerbara, men många tillstånd, t.ex. sicklecellsjukdom, minskar deras elasticitet, vilket gör dem mindre deformerbara. Röda blodkroppar med minskad deformbarhet har ett ökande flödeshinder, vilket leder till ökad aggregering av röda blodkroppar och minskad syremättnad, vilket kan leda till ytterligare komplikationer. Närvaron av celler med minskad deformbarhet, vilket är fallet vid sicklecellsjukdom, tenderar att hämma bildandet av plasmalager och genom att mäta viskoelasticiteten kan graden av hämning kvantifieras.
HistorikRedigera
I tidiga teoretiska arbeten behandlades blodet som en icke-Newtonsk viskös vätska. I de första studierna hade man utvärderat blod under stationärt flöde och senare med hjälp av oscillerande flöde. Professor George B. Thurston vid University of Texas presenterade först idén om att blod är viskoelastiskt 1972. De tidigare studierna som undersökte blod i jämnt flöde visade försumbara elastiska egenskaper eftersom den elastiska regimen lagras i blodet när flödet startar och därför är dess närvaro dold när flödet når ett jämnt tillstånd. I de tidiga studierna användes de egenskaper som konstaterades i ett stabilt flöde för att härleda egenskaper för instabila flödessituationer. Framsteg inom medicinska förfaranden och apparater krävde en bättre förståelse av blodets mekaniska egenskaper.