A vízelaszticitás az emberi vér olyan tulajdonsága, amely elsősorban a vörösvértestek deformációjában tárolt rugalmas energiának köszönhető, amikor a szív átpumpálja a vért a testen. A szív által a vérbe átadott energia részben a rugalmas szerkezetben tárolódik, másik része a viszkozitás révén disszipálódik, a fennmaradó energiát pedig a vér kinetikus mozgása tárolja. Ha figyelembe vesszük a szív pulzálását, világosan láthatóvá válik egy rugalmas rezsim. Kimutatták, hogy a vér tisztán viszkózus folyadékként való korábbi felfogása nem volt megfelelő, mivel a vér nem közönséges folyadék. A vért pontosabban rugalmas sejtek fluidizált szuszpenziójaként (vagy szolként) lehet leírni.
A vörösvértestek a vér térfogatának mintegy felét foglalják el, és rugalmas tulajdonságokkal rendelkeznek. Ez a rugalmas tulajdonság a legnagyobb mértékben hozzájárul a vér viszkoelasztikus viselkedéséhez. A vörösvértestek nagy térfogatszázaléka normál hematokritszint mellett kevés helyet hagy a sejtek mozgásának és deformációjának anélkül, hogy kölcsönhatásba lépnének egy szomszédos sejttel. Számítások kimutatták, hogy a vörösvértestek maximális térfogatszázalékának deformáció nélküli aránya 58%, ami a normálisan előforduló szintek tartományában van. A vörösvértestek közötti korlátozott tér miatt nyilvánvaló, hogy a vér áramlásához a sejtek közötti jelentős kölcsönhatás kulcsszerepet játszik. Ez a kölcsönhatás és a sejtek aggregálódási hajlama nagyban hozzájárul a vér viszkoelasztikus viselkedéséhez. A vörösvérsejtek deformációja és aggregációja az elrendeződés és orientáció áramlás okozta változásaival is párosul, ami a viszkoelasztikus viselkedés harmadik fő tényezője. A vér viszkoelasztikus tulajdonságaihoz hozzájáruló egyéb tényezők a plazma viszkozitása, a plazma összetétele, a hőmérséklet és az áramlási sebesség vagy nyírási sebesség. Ezek a tényezők együttesen teszik az emberi vért viszkoelasztikus, nem-newtoni és tixotróp vérré.
Amikor a vörösvértestek nyugalmi állapotban vagy nagyon kis nyírási sebességnél vannak, hajlamosak aggregálódni és energetikailag kedvező módon egymásra rakódni. A vonzást a sejtek felszínén lévő töltött csoportoknak, valamint a fibrinogén és a globulinok jelenlétének tulajdonítják. Ez az aggregált konfiguráció a sejteknek a legkisebb deformációval járó elrendeződése. Nagyon alacsony nyírási sebességek esetén a vér viszkoelasztikus tulajdonságát az aggregáció uralja, és a sejtek deformálhatósága viszonylag jelentéktelen. A nyírási sebesség növekedésével az aggregátumok mérete csökkenni kezd. A nyírási sebesség további növekedésével a sejtek átrendeződnek és orientálódnak, hogy csatornákat biztosítsanak a plazma áthaladásához és a sejtek csúszásához. Ebben az alacsony és közepes nyírási sebességtartományban a sejtek a szomszédos sejtekhez képest meginognak, lehetővé téve az áramlást. Az aggregációs tulajdonságok hatása a viszkoelaszticitásra csökken, és a vörösvérsejtek deformálhatóságának hatása kezd növekedni. Ahogy a nyírási sebesség nagy lesz, a vörösvértestek megnyúlnak vagy deformálódnak, és igazodnak az áramláshoz. Sejtrétegek alakulnak ki, amelyeket plazma választ el egymástól, és az áramlás most már a plazmarétegeken csúszó sejtrétegeknek tulajdonítható. A sejtréteg lehetővé teszi a vér könnyebb áramlását, és így csökken a viszkozitás és a rugalmasság. A vér viszkoelaszticitását a vörösvértestek deformálhatósága uralja.
Maxwell-modellSzerkesztés
A Maxwell-modell a Maxwell-folyadékokra vagy a Maxwell-anyagra vonatkozik. Az anyag a Maxwell modellben egy folyadék, ami azt jelenti, hogy tiszteletben tartja a konzervatív egyenletek folytonossági tulajdonságait : A folyadékok az anyagfázisok egy részhalmaza, és magukban foglalják a folyadékokat, a gázokat, a plazmákat és bizonyos mértékig a plasztikus szilárd anyagokat. A Maxwell-modell a viszkoelaszticitás helyi konzervatív értékeinek becslésére szolgál a modell integrális térfogatának globális mértékével, amelyet különböző áramlási helyzetekre lehet átültetni. A vér egy összetett anyag, ahol a különböző sejtek, mint például a vörösvértestek, diszkontinuusan helyezkednek el a plazmában. Méretük és alakjuk is szabálytalan, mivel nem tökéletes gömbök. A vértérfogat alakját tovább bonyolítja, hogy a vörösvérsejtek nem azonos módon oszlanak el a vérminta térfogatában, mert a sebességgradiensek irányában a legnagyobb sebességű területek felé vándorolnak, a Fåhræus-Lindqvist-effektus híres ábrázolása szerint, a Thurston által leírt hüvely- vagy dugóáramlásokban aggregálódnak vagy elkülönülnek. Jellemzően az alább leírt Maxwell-modell egyenletesen tekinti az anyagot (egyenletes kék szín) a térfogatban (kék színben) mindenütt tökéletesen eloszló részecskefolyadéknak, de Thurston feltárja, hogy a vörösvértestek csomagjai, dugók, inkább a nagy sebességű tartományban vannak jelen, ha y a Maxwell-modell ábráján a magasság iránya, (y~H) és az alacsonyabb sebességű területen (y~0) van egy szabad sejtréteg, ami azt jelenti, hogy a Maxwell-modell szerint deformálódó plazmafolyadék fázis a Maxwell-modell szerint feszült belső bélések után, amelyek teljesen kikerülnek az analitikus modellből Maxwell által.
Egy folyadék a Maxwell-modellben elméletileg pontosan hasonlóan viselkedik bármely más áramlási geometriában, például csövekben, forgó cellákban vagy nyugalmi állapotban. A gyakorlatban azonban a vér tulajdonságai a geometriától függően változnak, és a vérről kiderült, hogy nem megfelelő anyag a köznapi értelemben vett folyadékként való tanulmányozásra. Így a Maxwell-modell olyan tendenciákat ad, amelyeket a valós helyzetben ki kell egészíteni, majd egy edényben a Thurston-modell követi a sejtek eloszlását a köpeny- és dugóáramlásban.
Ha egy kis kocka alakú vérmennyiséget veszünk figyelembe, amelyre a szív pumpálása és a határokból eredő nyíróerők hatnak. A kocka alakváltozásának 2 összetevője lesz:
- Elasztikus deformáció, amely visszaállítható, és a vér szerkezetében tárolódik.
- A csúszás, amely folyamatos viszkózus energia bevitelével jár.
Az erő megszüntetésekor a kocka részben visszaállna. A rugalmas deformáció megfordul, de a csúszás nem. Ez megmagyarázza, hogy a rugalmas rész miért csak instacionárius áramlásban észlelhető. Állandósult áramlásban a csúszás tovább növekszik, és az időben nem változó erő mérései elhanyagolják a rugalmasság hozzájárulását.

Az 1. ábra segítségével kiszámíthatók a következő, a vér értékeléséhez szükséges paraméterek, amikor egy erőt kifejtünk.
Nyírófeszültség: τ = F A {\displaystyle \tau ={\frac {F}{A}}}

Nyírófeszültség: γ = D H {\displaystyle \gamma ={\frac {D}{H}}}

Nyírási sebesség: γ ˙ = V H {\displaystyle {\dot {\gamma }}={\frac {V}{H}}}}

A szív pulzációjának szimulálására szinuszos időben változó áramlást használunk. Az időben változó áramlásnak kitett viszkoelasztikus anyag fázisváltozást fog eredményezni τ {\displaystyle \tau }

és γ {\displaystyle \gamma }

által reprezentált ϕ {\displaystyle \phi }
között.

. Ha ϕ = 0 {\displaystyle \phi =0}

, az anyag tisztán rugalmas, mert a feszültség és az alakváltozás fázisban van, így az egyiknek a másik által okozott válasza azonnali. Ha ϕ {\displaystyle \phi }
= 90°, az anyag tisztán viszkózus, mert az alakváltozás 90 fokkal elmarad a feszültségtől. Egy viszkoelasztikus anyag valahol 0 és 90 fok között lesz.
A szinuszos időbeli változás e i ω t {\displaystyle e^{i\omega t}} arányos.

. Ezért a feszültség, az alakváltozás és a nyírási sebesség közötti méret- és fázisviszonyokat ezzel az összefüggéssel és egy radiánfrekvenciával, ω = 2 π f {\displaystyle \omega =2\pi f}

volt f {\displaystyle f}

a frekvencia hertzben kifejezve. Nyírófeszültség: τ ∗ = τ e – i ϕ {\displaystyle \tau ^{*}=\tau e^{-i\phi }}

Nyírófeszültség: γ ∗ = γ e – i π 2 {\displaystyle \gamma ^{*}=\gamma e^{-i{\frac {\pi }{2}}}}

Nyírási sebesség: γ ˙ ∗ = γ ˙ e – i 0 {\displaystyle {\dot {\dot {\gamma }}^{*}={\dot {\gamma }}e^{-i0}}}

A komplex nyírófeszültség összetevői a következőképpen írhatók fel:
τ ∗ = τ ′ – i τ ″ {\displaystyle \tau ^{*}=\tau ‘-i\tau ”}

Ha τ ′ {\displaystyle \tau ‘}

a viszkózus feszültség és τ ″ {\displaystyle \tau ”}

a rugalmas feszültség.A komplex viszkozitási együttható η ∗ {\displaystyle \eta ^{*}}

a komplex nyírófeszültség és a komplex nyírási sebesség hányadosaként határozható meg: η ∗ = τ ∗ γ ˙ ∗ = ( τ ′ γ ˙ + i τ ″ γ ˙ ) = η ′ + i η ″ {\displaystyle \eta ^{*}={\frac {\tau ^{*}}{{\dot {\gamma }}^{*}}}=({\frac {\tau ‘}{\dot {\gamma }}}+i{\frac {\tau ”}{\dot {\gamma }}})=\eta ‘+i\eta ”}

Hasonlóképpen a komplex dinamikus modulus G a komplex nyírófeszültség és a komplex nyírási alakváltozás hányadosaként kapható.
G = τ ∗ γ ∗ = ( τ ″ γ + i τ ′ γ ) {\displaystyle G={\frac {\tau ^{*}}{\gamma ^{*}}}}=({\frac {\tau ”}{\gamma }}+i{\frac {\tau ‘}{\gamma }})})}

Az egyenleteket az általános viszkoelasztikus kifejezésekre vonatkoztatva megkapjuk a tárolási modulust, G’, és a veszteségmodulust, G”.
G = G ′ + i G ″ {\displaystyle G=G’+iG”}

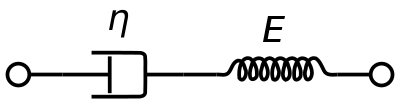
A vér viszkoelasztikus tulajdonságainak ábrázolására általában viszkoelasztikus Maxwell anyagmodellt használnak. Tisztán viszkózus csillapítót és egy tisztán rugalmas rugót használ sorba kapcsolva. E modell elemzése a komplex viszkozitást a dashpot-állandó és a rugóállandó függvényében adja meg.
η ∗ = η d a s h 1 + i ω ( η d a s h E s p r i n g ) = η ′ – i η ″ {\displaystyle \eta ^{*}={\frac {\eta _{dash}}{1+i\omega ({\frac {\eta _{dash}}{E_{spring}}})}}=\eta ‘-i\eta ”} }

Oldroyd-B modellEdit
A vér viszkoelaszticitásának egyik leggyakrabban használt konstitutív modellje az Oldroyd-B modell. Az Oldroyd-B nem-newtoni modellnek számos változata létezik, amelyek a vörösvérsejtek aggregációja és diszperziója miatt alacsony nyírási sebességnél fellépő nyírási vékonyodási viselkedést jellemzik. Itt egy háromdimenziós Oldroyd-B modellt vizsgálunk, amelyet az impulzusegyenlettel és a teljes feszültségtenzorral kapcsolunk össze. Nem newtoni áramlást alkalmazunk, amely biztosítja, hogy a vér μ ( h , d ) {\displaystyle \mu (h,d)} viszkozitása

az érátmérő d és a hematokrit h függvénye. Az Oldroyd-B modellben a B nyírófeszültség-tenzor és az A orientációs feszültség-tenzor közötti kapcsolat a következő:
S + γ = μ ( h , d ) – g A + C 1 ( g A – C 2 I μ ( h , d ) 2 ) {\displaystyle S+\gamma \left=\mu (h,d)\left-gA+C_{1}\left(gA-{\frac {C_{2}I}{\mu (h,d)^{2}}}}\right)}

ahol D/Dt az anyagi derivált, V a folyadék sebessége, C1, C2, g, γ {\displaystyle \gamma }
konstansok. S és B a következőképpen van definiálva: S = μ B + g A {\displaystyle S=\\mu B+gA}

B = Δ V + ( Δ V ) T {\displaystyle B=\Delta V+(\Delta V)^{T}}

A vörösvértestek viszkoelaszticitásaSzerkesztés
A vörösvértestek mind a véráramlás, mind az érfalak részéről intenzív mechanikai stimulációnak vannak kitéve, és reológiai tulajdonságaik fontosak a mikrokeringésben betöltött biológiai funkcióik hatékony ellátása szempontjából. Kimutatták, hogy a vörösvértestek önmagukban viszkoelasztikus tulajdonságokkal rendelkeznek. A vörösvértestek mechanikai tulajdonságainak feltárására számos módszert alkalmaznak, mint például:
- mikropipettás aspiráció
- mikrobehúzás
- optikai csipesz
- nagyfrekvenciás elektromos deformációs vizsgálatok
Ezek a módszerek a vörösvértestek deformálhatóságának jellemzésére dolgoztak a nyírási, hajlítási, területtágulási modulusok és relaxációs idők tekintetében. A viszkoelasztikus tulajdonságok feltárására azonban nem voltak képesek. Más technikákat is alkalmaztak, például fotoakusztikus méréseket. Ez a technika egyimpulzusos lézersugárral fotoakusztikus jelet hoz létre a szövetekben, és a jel lecsengési idejét mérik. A lineáris viszkoelaszticitás elméletének megfelelően a lecsengési idő megegyezik a viszkozitás-elaszticitás aránnyal, és így a vörösvértestek viszkoelaszticitási jellemzőit meg lehetett kapni.
A viszkoelaszticitás értékelésére használt másik kísérleti technika a sejtek felületére ragasztott ferromágneses gyöngyök használatából állt. Ezután a mágneses gyöngyre erőket alkalmaztak optikai mágneses csavaró citometria segítségével, ami lehetővé tette a kutatók számára a vörösvértestek időfüggő válaszainak feltárását.
T s ( t ) {\displaystyle T_{s}(t)}

az egységnyi gyöngy térfogatra jutó mechanikai nyomaték (a feszültség mértékegysége), és a következővel adódik: T s ( t ) = c H cos θ {\displaystyle T_{s}(t)=cH\cos \theta }

ahol H az alkalmazott mágneses csavarási tér, θ {\displaystyle {\theta }}

a gyöngy mágneses momentumának az eredeti mágnesezési irányhoz viszonyított szöge, c pedig a gyöngyállandó, amelyet a gyöngy ismert viszkozitású folyadékba helyezésével és csavarási tér alkalmazásával végzett kísérletekből találunk.
A G komplex dinamikus modulus használható az oszcilláló feszültség és az alakváltozás közötti összefüggések ábrázolására:
G = G ′ + i G ″ {\displaystyle G=G’+iG”}
ahol G ′ {\displaystyle G’}

a tárolási modulus és G ″ {\displaystyle G”}

a veszteségmodulus: G ′ = σ 0 ε 0 cos ϕ {\displaystyle G’={\frac {\sigma _{0}}{\varepsilon _{0}}}}\cos \phi }

G ″ = σ 0 ε 0 sin ϕ {\displaystyle G”={\frac {\sigma _{0}}{\varepsilon _{0}}}\sin \phi }

hol σ 0 {\displaystyle {\sigma _{0}}

és ε 0 {\displaystyle \varepsilon _{0}}

a feszültség és a nyúlás amplitúdója, és ϕ {\displaystyle \phi }
a köztük lévő fáziseltolódás.

A fenti összefüggésekből a komplex modulus összetevői egy olyan hurokból határozhatók meg, amely a nyomatékváltozás és az időváltozás összehasonlításával jön létre, amely grafikus ábrázolás esetén egy hurkot alkot. A T s ( t ) {\displaystyle T_{s}(t)} határai
– d(t) hurok és a T s ( t ) {\displaystyle T_{s}(t)} által határolt A terület.
– d(t) hurok, amely a ciklusonkénti energia disszipációját jelenti, a számítások során használjuk. A fázisszög ϕ
, a tárolási modulus G’ és a veszteség modulus G ezután a következő lesz: ϕ = sin – 1 4 A π Δ T s Δ d {\displaystyle \phi =\sin ^{-1}{\frac {4A}{\pi \Delta T_{s}\Delta d}}}

G ′ = Δ T s Δ d cos ϕ {\displaystyle G’={\frac {\Delta T_{s}}{\Delta d}}\cos \phi }\cos \phi }

G ″ = Δ T s Δ d sin ϕ = 4 A π ω Δ d 2 {\displaystyle G”={\frac {\Delta T_{s}}{\Delta d}}\sin \phi ={\frac {4A}{\pi \omega \Delta d^{2}}}}

ahol d az elmozdulás.
A 3. ábrán látható hiszterézis a vörösvértestekben jelenlévő viszkoelaszticitást reprezentálja. Nem világos, hogy ez a membrán molekuláris ingadozásaival vagy az intracelluláris ATP-koncentráció által szabályozott metabolikus aktivitással függ-e össze. További kutatásokra van szükség e kölcsönhatások teljes feltárásához és a vörösvértestek mögöttes viszkoelasztikus deformációs jellemzőinek megvilágításához.
A vérerek hatásaiSzerkesztés
A vér in vivo viszkoelasztikus viselkedésének vizsgálatakor figyelembe kell venni az artériák, kapillárisok és vénák hatásait is. A vér viszkozitása elsősorban a nagyobb artériákban befolyásolja az áramlást, míg a rugalmasság, amely a vörösvértestek rugalmas deformálhatóságában rejlik, elsősorban az arteriolákban és a kapillárisokban van hatással. Az artériás falakban történő hullámterjedés, a helyi hemodinamika és a fali nyírófeszültség-gradiens megértése fontos a szív- és érrendszeri működés mechanizmusainak megértéséhez. Az artériák falai anizotrópok és heterogének, különböző biomechanikai jellemzőkkel rendelkező rétegekből állnak, ami nagyon megnehezíti az artériák által a véráramláshoz hozzájáruló mechanikai hatások megértését.
A jobb megértés orvosi okaiSzerkesztés
A vér viszkoelasztikus tulajdonságainak tanulmányozásának fontossága orvosi szempontból nyilvánvalóvá válik. A szív- és érrendszeri protézisek, például szívbillentyűk és vérszivattyúk fejlesztésével szükségessé válik a pulzáló véráramlás megértése összetett geometriákban. Néhány konkrét példa a vér viszkoelaszticitásának hatása és annak következményei a pulzáló vérszivattyúk vizsgálatára. Erős összefüggéseket dokumentáltak a vér viszkoelaszticitása és a regionális és globális agyi véráramlás között a kardiopulmonális bypass alatt.
Ez vezetett a véranalógia kifejlesztéséhez is a protézisek tanulmányozása és tesztelése érdekében. A glicerin és víz klasszikus analógja jól reprezentálja a viszkozitást és a tehetetlenségi hatásokat, de nem rendelkezik a valódi vér rugalmas tulajdonságaival. Az egyik ilyen véranalóg a xantángumi és glicerin vizes oldata, amelyet úgy fejlesztettek ki, hogy megfeleljen a vér komplex viszkozitásának viszkózus és rugalmas összetevőinek egyaránt.
A normál vörösvértestek deformálhatók, de számos állapot, például a sarlósejtes betegség csökkenti a rugalmasságukat, ami miatt kevésbé deformálhatók. A csökkent deformálhatóságú vörösvértesteknek egyre nagyobb az áramlási akadálya, ami a vörösvértestek aggregációjának növekedéséhez és az oxigéntelítettség csökkenéséhez vezet, ami további szövődményekhez vezethet. A csökkent deformálhatóságú sejtek jelenléte, mint a sarlósejtes betegségben, hajlamos gátolni a plazmarétegek kialakulását, és a viszkoelaszticitás mérésével a gátlás mértéke számszerűsíthető.
ElőzményekSzerkesztés
A korai elméleti munkákban a vért nemnewtoni viszkózus folyadékként kezelték. A kezdeti tanulmányok a vért egyenletes áramlás során, később pedig oszcilláló áramlással értékelték. A Texasi Egyetem professzora, George B. Thurston 1972-ben mutatta be először a vér viszkoelasztikus voltának gondolatát. A korábbi tanulmányok, amelyek a vért állandósult áramlásban vizsgálták, elhanyagolható rugalmas tulajdonságokat mutattak, mivel a rugalmas rendszer az áramlás megindulása során tárolódik a vérben, és így jelenléte rejtve marad, amikor az áramlás eléri az állandósult állapotot. A korai tanulmányok az állandósult áramlásban talált tulajdonságokat használták fel az instacionárius áramlási helyzetekre vonatkozó tulajdonságok levezetésére. Az orvosi eljárások és eszközök fejlődése megkövetelte a vér mechanikai tulajdonságainak jobb megértését.