Visco-elasticiteit is een eigenschap van menselijk bloed die voornamelijk is toe te schrijven aan de elastische energie die is opgeslagen in de vervorming van rode bloedcellen wanneer het hart het bloed door het lichaam pompt. De energie die door het hart aan het bloed wordt overgedragen, wordt gedeeltelijk opgeslagen in de elastische structuur, een ander deel wordt afgevoerd door viscositeit, en de resterende energie wordt opgeslagen in de kinetische beweging van het bloed. Wanneer rekening wordt gehouden met de pulsatie van het hart, wordt een elastisch regime duidelijk. Aangetoond is dat het vroegere concept van bloed als een zuiver viskeuze vloeistof niet voldeed, aangezien bloed geen gewone vloeistof is. Bloed kan nauwkeuriger worden omschreven als een vloeibare suspensie van elastische cellen (of een sol).
De rode bloedcellen nemen ongeveer de helft van het bloedvolume in en bezitten elastische eigenschappen. Deze elastische eigenschap levert de grootste bijdrage aan het visco-elastische gedrag van bloed. Het grote volumepercentage van rode bloedcellen bij een normaal hematocrietgehalte laat weinig ruimte voor celbeweging en vervorming zonder interactie met een naburige cel. Berekeningen hebben aangetoond dat het maximale volumepercentage van rode bloedcellen zonder vervorming 58% bedraagt, hetgeen in het bereik ligt van normaal voorkomende niveaus. Door de beperkte ruimte tussen de rode bloedcellen is het duidelijk dat voor de doorstroming van het bloed een significante interactie tussen cellen een belangrijke rol zal spelen. Deze interactie en de neiging van cellen om samen te klonteren dragen in belangrijke mate bij tot het visco-elastische gedrag van bloed. De vervorming en aggregatie van rode bloedcellen is ook gekoppeld aan stromingsgeïnduceerde veranderingen in de ordening en oriëntatie als een derde belangrijke factor in het visco-elastische gedrag. Andere factoren die bijdragen tot de visco-elastische eigenschappen van bloed zijn de plasmaviscositeit, de samenstelling van het plasma, de temperatuur en de stromingssnelheid of afschuifsnelheid. Samen maken deze factoren menselijk bloed visco-elastisch, niet-Newtoniaans en thixotroop.
Wanneer de rode cellen in rust zijn of bij zeer kleine afschuifsnelheden, hebben zij de neiging om te aggregeren en op elkaar te stapelen op een energetisch gunstige manier. De aantrekkingskracht wordt toegeschreven aan geladen groepen op het oppervlak van de cellen en aan de aanwezigheid van fibrinogeen en globulinen. Deze geaggregeerde configuratie is een rangschikking van cellen met de minste vervorming. Bij zeer lage afschuifsnelheden wordt de visco-elastische eigenschap van bloed gedomineerd door de aggregatie en is de celvervormbaarheid relatief onbeduidend. Naarmate de afschuifsnelheid toeneemt, begint de grootte van de aggregaten af te nemen. Bij een verdere toename van de afschuifsnelheid zullen de cellen zich herschikken en oriënteren om kanalen te verschaffen waarlangs het plasma kan passeren en waarlangs de cellen kunnen glijden. In deze lage tot gemiddelde afschuifsnelheid bereik, de cellen wiebelen ten opzichte van de naburige cellen waardoor stroming. De invloed van aggregatie-eigenschappen op de visco-elasticiteit neemt af en de invloed van vervormbaarheid van de rode cel begint toe te nemen. Als de afschuifsnelheden groot worden, zullen de rode bloedcellen uitrekken of vervormen en zich met de stroming uitlijnen. Er worden cellagen gevormd, gescheiden door plasma, en de stroming wordt nu toegeschreven aan lagen cellen die over lagen plasma glijden. Door de cellaag kan het bloed gemakkelijker stromen en als zodanig is er een verminderde viscositeit en verminderde elasticiteit. De visco-elasticiteit van het bloed wordt gedomineerd door de vervormbaarheid van de rode bloedcellen.
Maxwell modelEdit
Maxwell Model betreft Maxwell vloeistoffen of Maxwell materiaal. Het materiaal in het Maxwell Model is een vloeistof, wat betekent dat het de continuïteitseigenschappen voor conservatieve vergelijkingen respecteert: Vloeistoffen zijn een deelverzameling van de fasen van materie en omvatten vloeistoffen, gassen, plasma’s en, tot op zekere hoogte, plastische vaste stoffen. Het Maxwell-model is gemaakt om lokale conservatieve waarden van visco-elasticiteit te schatten door een globale maat in het integrale volume van het model om te zetten naar verschillende stromingssituaties. Bloed is een complex materiaal waarin verschillende cellen zoals rode bloedcellen discontinu zijn in plasma. Hun grootte en vorm zijn ook onregelmatig omdat het geen perfecte bollen zijn. Wat de vorm van het bloedvolume nog ingewikkelder maakt, is dat de rode cellen niet identiek verdeeld zijn in een bloedmonster, omdat zij met snelheidsgradiënten in de richting van de gebieden met de hoogste snelheid migreren, wat de beroemde voorstelling van het Fåhræus-Lindqvist effect is, zich aggregeren of scheiden in mantel- of stopstromingen, zoals beschreven door Thurston. Typisch, beschouwt het hieronder beschreven Maxwell Model uniform het materiaal (uniforme blauwe kleur) als een perfecte verdeelde deeltjesvloeistof overal in het volume (in blauw) maar Thurston openbaart dat de pakketten van rode cellen, stekkers, meer aanwezig in het hoge snelheidsgebied zijn, als y de hoogterichting in het Maxwell modelcijfer is, (y~H) en er is een vrije cellenlaag in het lagere snelheidsgebied (y~0) wat betekent de plasmavloeistoffase die onder Maxwell Model vervormt is gespannen na binnenvoeringen die volledig aan het analytische model door Maxwell ontsnappen.
In theorie gedraagt een vloeistof in een Maxwell Model zich precies zo in elke andere stromingsgeometrie zoals pijpen, roterende cellen of in rusttoestand. Maar in de praktijk variëren de eigenschappen van het bloed met de geometrie en is bloed een ongeschikt materiaal gebleken om als een vloeistof in gewone zin te worden bestudeerd. Dus geeft het Maxwell Model tendensen die in de werkelijke situatie moeten worden aangevuld, gevolgd door het Thurston model in een vat met betrekking tot de verdeling van cellen in mantel- en stopstromen.
Als een klein kubusvormig volume bloed wordt beschouwd, met krachten die erop werken door het pompen van het hart en afschuifkrachten van de grenzen. De vormverandering van de kubus zal 2 componenten hebben:
- Elastische vervorming die kan worden hersteld en is opgeslagen in de structuur van het bloed.
- Slippage die gepaard gaat met een voortdurende toevoer van viskeuze energie.
Wanneer de kracht wordt weggenomen, zou de kubus zich gedeeltelijk herstellen. De elastische vervorming is omgekeerd, maar de slip niet. Dit verklaart waarom het elastische deel alleen merkbaar is bij onbestendige stroming. Bij gestage stroming zal de slip blijven toenemen en zullen de metingen van niet in de tijd variërende kracht de bijdragen van de elasticiteit verwaarlozen.

Figuur 1 kan worden gebruikt om de volgende parameters te berekenen die nodig zijn voor de evaluatie van bloed wanneer een kracht wordt uitgeoefend.
Afschuifspanning: τ = F A {\displaystyle \tau ={\frac {F}{A}}}

Afschuifspanning: γ = D H {\displaystyle \gamma ={\frac {D}{H}}}

Schuifsnelheid: γ ˙ = V H {{\displaystyle {\gamma}}={\frac {V}{H}}}

Een sinusoïdale tijdsvariërende stroming wordt gebruikt om de pulsatie van een hart te simuleren. Een visco-elastisch materiaal dat wordt onderworpen aan een tijdsvariërende stroming zal resulteren in een fasevariatie tussen τ {\displaystyle \tau }

en γ {\displaystyle \gamma }

vertegenwoordigd door ϕ {\displaystyle \phi }

. Als ϕ = 0 {\displaystyle \phi =0}

, is het materiaal zuiver elastisch omdat spanning en rek in fase zijn, zodat de reactie van de een veroorzaakt door de ander onmiddellijk is. Als ϕ {\displaystyle \phi }
= 90°, is het materiaal zuiver viskeus omdat de rek 90 graden achterloopt op de spanning. Een visco-elastisch materiaal zal ergens tussen 0 en 90 graden liggen.
De sinusvormige tijdvariatie is evenredig met e i ω t {\displaystyle e^{i\omega t}}

. Daarom worden de grootte- en faserelatie tussen de spanning, rek en afschuifsnelheid beschreven met behulp van deze relatie en een radiale frequentie, ω = 2 π f {Displaystyle \omega =2 f}

waren f {\displaystyle f}

de frequentie in Hertz is. Afschuifspanning: τ ∗ = τ e – i ϕ {\displaystyle \tau ^{*}=\tau e^{-i\phi }}

Afschuifspanning: γ ∗ = γ e – i π 2 {Displaystyle \gamma ^{*}=\gamma e^{-i{\frac {pi }{2}}}}

Schuifsnelheid: γ ˙ ∗ = γ ˙ e – i 0 {\displaystyle {{\dot {\gamma}}^{*}={\dot {\gamma }}e^{-i0}}

De componenten van de complexe afschuifspanning kunnen worden geschreven als:
τ ∗ = τ ′ – i τ ″ {\displaystyle \tau ^{*}=\tau ‘-i”}

Waar τ ′ {Displaystyle \tau ‘}

de viskeuze spanning is en τ ″ {\displaystyle \tau ”}

is de elastische spanning.De complexe viscositeitscoëfficiënt η ∗ {\displaystyle \eta ^{*}}

kan worden gevonden door de verhouding te nemen tussen de complexe afschuifspanning en de complexe afschuifsnelheid: η ∗ = τ ∗ γ ˙ ∗ = ( τ ′ γ ˙ + i τ ″ γ ˙ ) = η ′ + i η ″ {{\displaystyle γeta ^{*}={\frac {\tau ^{*}}{\dot {\gamma }}^{*}}=({\frac {\tau ‘}{\dot {\gamma }}+i{\frac {\tau ”}{\dot {\gamma }})={\tau ‘+i\tau ”}}+i{\frac {\tau ”}}

Op soortgelijke wijze kan de complexe dynamische modulus G worden verkregen door de verhouding te nemen tussen de complexe schuifspanning en de complexe schuifspanning.
G = τ ∗ γ ∗ = ( τ ″ γ + i τ ′ γ ) {\displaystyle G={\frac {\tau ^{*}}{\gamma ^{*}}}=({\frac {\tau ”}{\gamma }}+i{\frac {\tau ‘}{\gamma }})}

Door de vergelijkingen te relateren aan gebruikelijke visco-elastische termen krijgen we de opslagmodulus, G’, en de verliesmodulus, G”.
G = G ′ + i G ″ {Displaystyle G=G’+iG”}

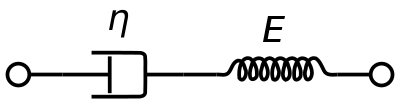
Een visco-elastisch Maxwell-materiaalmodel wordt algemeen gebruikt om de visco-elastische eigenschappen van bloed weer te geven. Het maakt gebruik van een zuiver viskeuze demper en een zuiver elastische veer die in serie zijn geschakeld. Analyse van dit model geeft de complexe viscositeit in termen van de dashpotconstante en de veerconstante.
η ∗ = η d a s h 1 + i ω ( η d a s h E s p r i n g ) = η ′ – i η ″ {\displaystyle \eta ^{*}={\frac {\eta _{dash}}{1+i\omega ({\frac {\eta _{dash}}{E_{spring}}})}}= η ‘-i\eta ”}.

Oldroyd-B modelEdit
Een van de meest gebruikte constitutieve modellen voor de visco-elasticiteit van bloed is het Oldroyd-B model. Er zijn verschillende variaties van het niet-Newtoniaanse Oldroyd-B model dat afschuifverdunnend gedrag karakteriseert ten gevolge van aggregatie en dispersie van rode bloedcellen bij lage afschuifsnelheden. Hier beschouwen wij een driedimensionaal Oldroyd-B model gekoppeld aan de impulsvergelijking en de totale spanningstensor. Er wordt gebruik gemaakt van een niet-Newtoniaanse stroming, zodat de viscositeit van bloed μ ( h , d ) {\displaystyle \mu (h,d)}

een functie is van de vaatdiameter d en het hematocriet h. In het Oldroyd-B model wordt het verband tussen de schuifspanningstensor B en de oriëntatiespanningstensor A gegeven door:
S + γ = μ ( h , d ) – g A + C 1 ( g A – C 2 I μ ( h , d ) 2 ) {Displaystyle S+gamma \left=gmu (h,d)\left-gA+C_{1}\left(gA-{\frac {C_{2}I}{mu (h,d)^{2}}}}

waarbij D/Dt de materiaalafgeleide is, V de snelheid van de vloeistof, C1, C2, g, γ {\displaystyle \gamma }
zijn constanten. S en B zijn als volgt gedefinieerd: S = μ B + g A {{Displaystyle S==mu B+gA}

B = Δ V + ( Δ V ) T {\displaystyle B=(\Delta V)^{T}}

Visco-elasticiteit van rode bloedcellenEdit
Rode bloedcellen worden intensief mechanisch geprikkeld door zowel de bloedstroom als de vaatwanden, en hun reologische eigenschappen zijn belangrijk voor hun effectiviteit bij het uitvoeren van hun biologische functies in de microcirculatie. Van rode bloedcellen zelf is aangetoond dat zij visco-elastische eigenschappen bezitten. Er zijn verschillende methoden gebruikt om de mechanische eigenschappen van rode bloedcellen te onderzoeken, zoals:
- micropipet aspiratie
- micro-indentatie
- optisch pincet
- hoge frequentie elektrische vervormingstesten
Deze methoden werkten om de vervormbaarheid van de rode bloedcel te karakteriseren in termen van de afschuiving, buiging, gebiedsuitbreiding moduli, en ontspanningstijden. Zij waren echter niet in staat om de visco-elastische eigenschappen te onderzoeken. Andere technieken zijn toegepast, zoals fotoakoestische metingen. Deze techniek maakt gebruik van een laserbundel met één enkele puls om een fotoakoestisch signaal in weefsels op te wekken en de vervaltijd van het signaal wordt gemeten. Volgens de theorie van de lineaire visco-elasticiteit is de vervaltijd gelijk aan de verhouding viscositeit-elasticiteit, zodat de visco-elasticiteitskenmerken van de rode bloedcellen kunnen worden verkregen.
Een andere experimentele techniek die is gebruikt om de visco-elasticiteit te evalueren, bestond uit het gebruik van ferromagnetismekorrels die aan het oppervlak van een cel waren gehecht. Vervolgens worden krachten uitgeoefend op de magnetische kraal met behulp van optische magnetische verdraaiingscytometrie, waardoor onderzoekers de tijdsafhankelijke reacties van rode bloedcellen konden onderzoeken.
T s ( t ) {\displaystyle T_{s}(t)}

is de mechanische torsie per eenheid kraalvolume (eenheden van spanning) en wordt gegeven door: T s ( t ) = c H cos θ {Displaystyle T_{s}(t)=cH cos θtheta }

waarbij H het toegepaste magnetische verdraaiingsveld is, θ {\displaystyle {\theta }}

de hoek is van het magnetische moment van de kraal ten opzichte van de oorspronkelijke magnetisatierichting, en c de kraalconstante die wordt gevonden door experimenten waarbij de kraal in een vloeistof met een bekende viscositeit wordt geplaatst en een verdraaiingsveld wordt toegepast.
De complexe dynamische modulus G kan worden gebruikt om de relaties tussen de oscillerende spanning en rek weer te geven:
G = G ′ + i G ″ {\displaystyle G=G’+iG”}
waarbij G ′ {\displaystyle G’}

de opslagmodulus is en G ″ {\displaystyle G”}

is de verliesmodulus: G ′ = σ 0 ε 0 cos ϕ {Displaystyle G’={\frac {\sigma _{0}}{varepsilon _{0}}}\cos \phi }

G ″ = σ 0 ε 0 sin ϕ {\displaystyle G”={\frac {sigma _{0}}{varepsilon _{0}}}sin \phi }

waar σ 0 {\displaystyle \sigma _{0}}

en ε 0 {\displaystyle \varepsilon _{0}}

zijn de amplitudes van spanning en rek en ϕ {\displaystyle \phi }
is de faseverschuiving tussen beide.

Uit bovenstaande relaties worden de componenten van de complexe modulus bepaald uit een lus die ontstaat door de verandering in het koppel te vergelijken met de verandering in tijd, die een lus vormt wanneer deze grafisch wordt weergegeven. De grenzen van T s ( t ) {Displaystyle T_{s}(t)}
– d(t) lus en het gebied, A, begrensd door de T s ( t ) {\displaystyle T_{s}(t)}
– d(t)-lus, die de energiedissipatie per cyclus weergeeft, worden in de berekeningen gebruikt. De fasehoek ϕ {\displaystyle \phi }
, opslagmodulus G’, en verliesmodulus G worden dan: ϕ = sin – 1 4 A π Δ T s Δ d {\displaystyle \phi =\sin ^{-1}{\frac {4A}{\pi \Delta T_{s}\Delta d}}}

G ′ = Δ T s Δ d cos ϕ {\displaystyle G’={\frac {\Delta T_{s}}{Delta d}}\cos \phi }

G ″ = Δ T s Δ d sin ϕ = 4 A π ω Δ d 2 {\displaystyle G”={\frac {\Delta T_{s}}{\Delta d}}\sin \phi ={\frac {4A}{\pi \omega \Delta d^{2}}}}

waarbij d de verplaatsing is.
De in figuur 3 getoonde hysterese vertegenwoordigt de in rode bloedcellen aanwezige visco-elasticiteit. Het is onduidelijk of dit verband houdt met membraan moleculaire fluctuaties of metabole activiteit gecontroleerd door intracellulaire concentraties van ATP. Verder onderzoek is nodig om deze interactie volledig te onderzoeken en licht te werpen op de onderliggende visco-elastische vervormingskarakteristieken van de rode bloedcellen.
Effecten van bloedvatenEdit
Bij het bekijken van visco-elastisch gedrag van bloed in vivo, is het noodzakelijk om ook de effecten van slagaders, haarvaten en aders in beschouwing te nemen. De viscositeit van bloed heeft een primaire invloed op de stroming in de grotere arteriën, terwijl de elasticiteit, die berust op de elastische vervormbaarheid van rode bloedcellen, een primaire invloed heeft in de arteriolen en de haarvaten. Inzicht in de golfvoortplanting in slagaderwanden, de lokale hemodynamica en de wandschuifspanningsgradiënt is belangrijk om de mechanismen van de cardiovasculaire functie te begrijpen. Slagaderwanden zijn anisotroop en heterogeen, samengesteld uit lagen met verschillende biomechanische eigenschappen, wat het begrijpen van de mechanische invloeden die slagaders bijdragen aan de bloedstroom zeer moeilijk maakt.
Medische redenen voor een beter begripEdit
Vanuit een medisch standpunt wordt het belang van het bestuderen van de visco-elastische eigenschappen van bloed duidelijk. Met de ontwikkeling van cardiovasculaire prothesen, zoals hartkleppen en bloedpompen, is het begrijpen van pulserende bloedstroming in complexe geometrieën vereist. Een paar specifieke voorbeelden zijn de effecten van de visco-elasticiteit van bloed en de implicaties daarvan voor het testen van een pulserende bloedpomp. Sterke correlaties tussen de visco-elasticiteit van bloed en de regionale en globale cerebrale bloedstroom tijdens cardiopulmonaire bypass zijn gedocumenteerd.
Dit heeft ook de weg gebaand voor de ontwikkeling van een bloedanalogon om prothetische hulpmiddelen te bestuderen en te testen. De klassieke analoog van glycerine en water biedt een goede weergave van de viscositeit en inertie-effecten, maar mist de elastische eigenschappen van echt bloed. Eén zo’n bloedanalogon is een waterige oplossing van Xanthaangom en glycerine, ontwikkeld om zowel de viskeuze als de elastische componenten van de complexe viscositeit van bloed te evenaren.
Normale rode bloedcellen zijn vervormbaar, maar veel aandoeningen, zoals sikkelcelziekte, verminderen hun elasticiteit, waardoor ze minder vervormbaar zijn. Rode bloedcellen met verminderde vervormbaarheid hebben een toenemende stromingsweerstand, wat leidt tot een toename van de aggregatie van rode bloedcellen en een verlaging van de zuurstofverzadiging, wat tot verdere complicaties kan leiden. De aanwezigheid van cellen met verminderde vervormbaarheid, zoals het geval is bij sikkelcelziekte, heeft de neiging de vorming van plasmalagen te remmen en door de visco-elasticiteit te meten, kan de mate van remming worden gekwantificeerd.
GeschiedenisEdit
In vroege theoretische werken werd bloed behandeld als een niet-Newtoniaanse viskeuze vloeistof. De eerste studies evalueerden het bloed bij constante stroming en later bij oscillerende stroming. Professor George B. Thurston, van de Universiteit van Texas, kwam in 1972 met het idee dat bloed visco-elastisch is. De eerdere studies die naar bloed in constante stroming keken, toonden verwaarloosbare elastische eigenschappen, omdat het elastische regime in het bloed is opgeslagen tijdens het op gang komen van de stroming en de aanwezigheid ervan dus verborgen blijft wanneer een stroming een constante toestand bereikt. De vroege studies gebruikten de eigenschappen gevonden in stabiele stroming om eigenschappen af te leiden voor onstabiele stromingssituaties. Vooruitgang in medische procedures en apparatuur vereiste een beter begrip van de mechanische eigenschappen van bloed.