Viskoelastisuus on ihmisveren ominaisuus, joka johtuu ensisijaisesti elastisesta energiasta, joka varastoituu punasolujen muodonmuutoksiin, kun sydän pumppaa verta kehon läpi. Sydämen vereen siirtämä energia varastoituu osittain kimmoisaan rakenteeseen, toinen osa haihtuu viskositeetin vaikutuksesta, ja loput energiasta varastoituu veren kineettiseen liikkeeseen. Kun sydämen sykintä otetaan huomioon, elastinen järjestelmä tulee selvästi esiin. On osoitettu, että aiempi käsitys verestä puhtaasti viskoosisena nesteenä oli riittämätön, koska veri ei ole tavallinen neste. Verta voidaan tarkemmin kuvata elastisten solujen fluidisoituneena suspensiona (tai solina).
Punasolut vievät noin puolet veren tilavuudesta, ja niillä on elastisia ominaisuuksia. Tämä elastinen ominaisuus on suurin veren viskoelastiseen käyttäytymiseen vaikuttava tekijä. Punasolujen suuri tilavuusprosentti normaalilla hematokriittitasolla jättää vain vähän tilaa solujen liikkeelle ja muodonmuutokselle ilman vuorovaikutusta naapurisolun kanssa. Laskelmat ovat osoittaneet, että punasolujen suurin tilavuusprosentti ilman muodonmuutosta on 58 %, mikä on normaalisti esiintyvien tasojen alueella. Koska punasolujen välinen tila on rajallinen, on selvää, että veren virtauksen kannalta solujen välinen vuorovaikutus on avainasemassa. Tämä vuorovaikutus ja solujen taipumus aggregaatioon vaikuttavat merkittävästi veren viskoelastiseen käyttäytymiseen. Punasolujen muodonmuutos ja aggregaatio liittyy myös virtauksen aiheuttamiin muutoksiin niiden järjestyksessä ja orientaatiossa, mikä on kolmas merkittävä tekijä sen viskoelastisessa käyttäytymisessä. Muita veren viskoelastisiin ominaisuuksiin vaikuttavia tekijöitä ovat plasman viskositeetti, plasman koostumus, lämpötila ja virtausnopeus tai leikkausnopeus. Yhdessä nämä tekijät tekevät ihmisverestä viskoelastisen, ei-newtonilaisen ja tiksotrooppisen.
Kun punasolut ovat levossa tai hyvin pienillä leikkausnopeuksilla, niillä on taipumus aggregoitua ja pinoutua yhteen energeettisesti edullisella tavalla. Vetovoima johtuu solujen pinnalla olevista varautuneista ryhmistä sekä fibrinogeenin ja globuliinien läsnäolosta. Tämä aggregoitunut konfiguraatio on solujen järjestely, jossa on vähiten muodonmuutoksia. Hyvin alhaisilla leikkausnopeuksilla veren viskoelastista ominaisuutta hallitsee aggregaatio, ja solujen muodonmuutoskyky on suhteellisen merkityksetön. Leikkausnopeuden kasvaessa aggregaattien koko alkaa pienentyä. Kun leikkausnopeus kasvaa edelleen, solut järjestäytyvät uudelleen ja suuntautuvat siten, että plasma pääsee kulkemaan kanavien läpi ja solut liukuvat. Tällä alhaisen tai keskisuuren leikkausnopeuden alueella solut heiluvat naapurisoluihin nähden, mikä mahdollistaa virtauksen. Aggregaatio-ominaisuuksien vaikutus viskoelastisuuteen vähenee ja punasolujen muodonmuutoskyvyn vaikutus alkaa kasvaa. Kun leikkausnopeus kasvaa suureksi, punasolut venyvät tai deformoituvat ja mukautuvat virtauksen mukaan. Solukerroksia muodostuu plasman erottamina, ja virtaus johtuu nyt solukerrosten liukumisesta plasmakerrosten päällä. Solukerros helpottaa veren virtausta, jolloin viskositeetti ja elastisuus vähenevät. Veren viskoelastisuutta hallitsee punasolujen muodonmuutoskyky.
Maxwellin malliEdit
Maxwellin malli koskee Maxwellin nesteitä tai Maxwellin materiaalia. Materiaali Maxwell-mallissa on neste, mikä tarkoittaa, että se noudattaa jatkuvuusominaisuuksia konservatiivisten yhtälöiden osalta: Fluidit ovat aineen faasien osajoukko ja niihin kuuluvat nesteet, kaasut, plasmat ja jossain määrin myös plastiset kiinteät aineet. Maxwellin malli on tehty arvioimaan viskoelastisuuden paikallisia konservatiivisia arvoja globaalin toimenpiteen avulla mallin integraalitilavuudessa, joka voidaan siirtää erilaisiin virtaustilanteisiin. Veri on monimutkainen materiaali, jossa eri solut, kuten punasolut, ovat epäjatkuvia plasmassa. Myös niiden koko ja muoto ovat epäsäännöllisiä, koska ne eivät ole täydellisiä palloja. Veren tilavuuden muotoa mutkistaa lisäksi se, että punasolut eivät jakaudu identtisesti verinäytteen tilavuuteen, koska ne vaeltavat nopeusgradienttien avulla suurimman nopeuden alueille, jotka ovat kuuluisan Fåhræus-Lindqvist-ilmiön kuvauksen mukaisia, ja ne aggregoituvat tai erkaantuvat Thurstonin kuvaamissa vaippa- tai tulppavirtauksissa. Tyypillisesti alla kuvatussa Maxwell-mallissa tarkastellaan materiaalia (yhtenäinen sininen väri) yhtenäisesti täydellisesti jakautuneena hiukkasnesteenä kaikkialla tilavuudessa (sinisellä), mutta Thurston paljastaa, että punasolupaketteja, tulppia, on enemmän korkealla nopeusalueella, jos y on korkeussuunta Maxwellin mallin kuviossa (y~H) ja alemmalla nopeusalueella on vapaita soluja sisältävä kerros (y~0), mikä tarkoittaa sitä, että Maxwellin mallin mukaisesti muokkautuva plasman nestemäinen vaihe jännittyy sisempien vuorausten jälkeen, jotka karkaavat täydellisesti Maxwellin esittämästä analyyttisestä mallista.
Teoriassa Maxwellin mallissa oleva neste käyttäytyy täsmälleen samalla tavalla missä tahansa muussa virtausgeometriassa, kuten putkissa, pyörivissä kennoissa tai lepotilassa. Käytännössä veren ominaisuudet kuitenkin vaihtelevat geometrian mukaan, ja veri on osoittautunut riittämättömäksi materiaaliksi tutkittavaksi nesteenä yleisessä mielessä. Niinpä Maxwellin malli antaa suuntauksia, joita on täydennettävä todellisessa tilanteessa, jota seuraa Thurstonin malli verisuonessa koskien solujen jakautumista vaippa- ja tulppavirtauksissa.
Jos tarkastellaan pientä kuutiomaista veritilavuutta, johon kohdistuu sydämen pumppauksen aiheuttamia voimia ja rajapinnoilta tulevia leikkausvoimia. Kuution muodonmuutoksella on 2 komponenttia:
- Elastinen muodonmuutos, joka on palautuva ja joka on varastoitunut veren rakenteeseen.
- Liukuma, joka liittyy jatkuvaan viskoosienergian syöttöön.
Kun voima poistetaan, kuutio palautuisi osittain. Kimmoinen muodonmuutos kumoutuu, mutta liukuminen ei. Tämä selittää, miksi elastinen osuus on havaittavissa vain epävakaassa virtauksessa. Tasaisessa virtauksessa liukuma kasvaa edelleen, ja ajassa muuttumattoman voiman mittauksissa kimmoisuuden osuus jää huomiotta.

Kuvasta 1 voidaan laskea seuraavat parametrit, joita tarvitaan veren arvioimiseksi voimaa käytettäessä.
Leikkausjännitys: τ = F A {\displaystyle \tau ={\frac {F}{A}}}

Shear Strain: γ = D H {\displaystyle \gamma ={\frac {D}{H}}}

Leikkausnopeus: γ ˙ = V H {\displaystyle {\dot {\gamma }}={\frac {V}{H}}}

Sinimuotoista ajallisesti vaihtelevaa virtausta käytetään simuloimaan sydämen sykettä. Viskoelastinen materiaali, johon kohdistuu ajallisesti vaihteleva virtaus, johtaa vaihevaihteluun välillä τ {\displaystyle \tau }.

ja γ {\displaystyle \gamma } välillä.

edustettuna ϕ {\displaystyle \phi }

. Jos ϕ = 0 {\displaystyle \phi =0}

, materiaali on puhtaasti elastinen, koska jännitys ja venymä ovat faasissa, joten toisen aiheuttama vaste on välitön. Jos ϕ {\displaystyle \phi }
= 90°, materiaali on puhtaasti viskoosinen, koska venymä on 90 astetta jäljessä jännityksestä. Viskoelastinen materiaali on jossain 0 ja 90 asteen välillä.
Sinimuotoinen aikavaihtelu on verrannollinen e i ω t {\displaystyle e^{i\omega t}}

. Näin ollen jännityksen, muodonmuutoksen ja leikkausnopeuden välinen suuruus- ja vaihesuhde kuvataan käyttämällä tätä suhdetta ja radiaalista taajuutta ω = 2 π f {\displaystyle \omega =2\pi f}

olivat f {\displaystyle f}

on taajuus hertseinä. Leikkausjännitys: τ ∗ = τ e – i ϕ {\displaystyle \tau ^{*}=\tau e^{-i\phi }}

Shear Strain: γ ∗ = γ e – i π 2 {\displaystyle \gamma ^{*}=\gamma e^{-i{\frac {\pi }{2}}}}

Leikkausnopeus: γ ˙ ∗ = γ ˙ e – i 0 {\displaystyle {\dot {\gamma }}^{*}={\dot {\gamma }}e^{-i0}}}

Kompleksisen leikkausjännityksen komponentit voidaan kirjoittaa seuraavasti:
τ ∗ = τ ′ – i τ ″ {\displaystyle \tau ^{*}=\tau ’-i\tau ”}

Jossa τ ′ {\displaystyle \tau ’}

on viskoosijännitys ja τ ″ {\displaystyle \tau ”}

on elastinen jännitys.Kompleksinen viskositeettikerroin η ∗ {\displaystyle \eta ^{*}}

saadaan ottamalla kompleksisen leikkausjännityksen ja kompleksisen leikkausnopeuden suhde: η ∗ = τ ∗ γ ˙ ∗ = ( τ ′ γ ˙ + i τ ″ γ ˙ ) = η ′ + i η ″ {\displaystyle \eta ^{*}={\frac {\tau ^{*}}{{\dot {\gamma }}^{*}}}=({\frac {\tau ’}{\dot {\gamma }}}+i{\frac {\tau ”}{\dot {\gamma }}})=\eta ’+i\eta ”}

Vastaavasti kompleksinen dynaaminen moduuli G saadaan ottamalla kompleksisen leikkausjännityksen ja kompleksisen leikkausvenymän suhde.
G = τ ∗ γ ∗ = ( τ ″ γ + i τ ′ γ ) {\displaystyle G={\frac {\tau ^{*}}{\gamma ^{*}}}}=({\frac {\tau ”}{\gamma }}+i{\frac {\tau ”}{\gamma }})})

Suhteuttamalla yhtälöt yleisiin viskoelastisiin termeihin saadaan varastointimoduuli G’ ja häviömoduuli G”.
G = G ′ + i G ″ {\displaystyle G=G’+iG”}

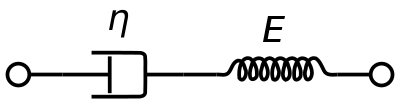
Viskoelastista Maxwellin materiaalimallia käytetään yleisesti veren viskoelastisten ominaisuuksien esittämiseen. Siinä käytetään sarjaan kytkettyjä puhtaasti viskoosia vaimenninta ja puhtaasti elastista jousta. Tämän mallin analyysi antaa kompleksisen viskositeetin dashpot-vakion ja jousivakion avulla.
η ∗ = η d a s h 1 + i ω ( η d a s h E s p r i n g ) = η ′ – i η ″ {\displaystyle {\displaystyle \eta ^{*}={\frac {\eta _{dash}}{1+i\omega ({\frac {\eta _{dash}}{E_{jousi}})}}=\eta ’-i\eta ”}}

Oldroyd-B-malliEdit
Yksi yleisimmin käytetyistä veren viskoelastisuuden konstitutiivisista malleista on Oldroyd-B-malli. Oldroyd-B:n ei-newtonilaisesta mallista on olemassa useita muunnelmia, jotka kuvaavat punasolujen aggregaatiosta ja dispersiosta johtuvaa leikkausohennuskäyttäytymistä alhaisella leikkausnopeudella. Tässä tarkastellaan kolmiulotteista Oldroyd-B-mallia, joka on yhdistetty momenttiyhtälöön ja kokonaisjännitystensoriin. Käytetään ei-newtonilaista virtausta, joka takaa, että veren viskositeetti μ ( h , d ) {\displaystyle \mu (h,d)}

on verisuonen halkaisijan d ja hematokriitin h funktio. Oldroyd-B-mallissa leikkausjännitystensorin B ja orientaatiojännitystensorin A välinen suhde saadaan seuraavasti:
S + γ = μ ( h , d ) – g A + C 1 ( g A – C 2 I μ ( h , d ) 2 ) {\displaystyle S+\gamma \left=\mu (h,d)\left-gA+C_{1}\left(gA-{\frac {C_{2}I}{\mu (h,d)^{2}}}\right)}

jossa D/Dt on aineen derivaatta, V on nesteen nopeus, C1, C2, g, γ {\displaystyle \gamma }
ovat vakioita. S ja B määritellään seuraavasti: S = μ B + g A {\displaystyle S=\mu B+gA}

B = Δ V + ( Δ V ) T {\displaystyle B=\Delta V+(\Delta V)^{T}}

Punasolujen viskoelastisuusEdit
Punasolut altistuvat voimakkaalle mekaaniselle stimulaatiolle sekä verenkierron että verisuonten seinämien taholta, ja niiden reologiset ominaisuudet ovat tärkeitä niiden tehokkuuden kannalta niiden suorittaessa biologisia tehtäviään mikroverenkierrossa. Punasoluilla itsellään on osoitettu olevan viskoelastisia ominaisuuksia. Punasolujen mekaanisten ominaisuuksien tutkimiseen käytetään useita menetelmiä, kuten seuraavia:
- mikropipetin aspiraatio
- mikroindentaatio
- optiset pinsetit
- korkeataajuuksiset sähköiset muodonmuutoskokeet
Nämä menetelmät työskentelivät punaisten verisolujen muodonmuutosominaisuuksien karakterisoimiseksi leikkaus-, taivutus-, pinta-alalaajenemis- ja relaksaatioaikojen osalta. Niillä ei kuitenkaan pystytty tutkimaan viskoelastisia ominaisuuksia. On toteutettu muita tekniikoita, kuten fotoakustisia mittauksia. Tässä tekniikassa käytetään yhden pulssin lasersädettä fotoakustisen signaalin synnyttämiseksi kudoksissa, ja signaalin hajoamisaika mitataan. Lineaarisen viskoelastisuuden teorian mukaan hajoamisaika on yhtä suuri kuin viskositeetti-elastisuussuhde, joten punasolujen viskoelastisuusominaisuudet saatiin selville.
Toinen kokeellinen tekniikka, jota käytettiin viskoelastisuuden arvioimiseksi, koostui solujen pintaan liimattujen ferromagnetismihelmien käytöstä. Tämän jälkeen magneettihelmiin kohdistetaan voimia käyttämällä optista magneettista vääntösytometriaa, jonka avulla tutkijat pystyivät tutkimaan punasolujen ajasta riippuvia vasteita.
T s ( t ) {\displaystyle T_{s}(t)}

on mekaaninen vääntömomentti helmiäisen tilavuusyksikköä kohti (jännitysyksiköt) ja se saadaan kaavalla: T s ( t ) = c H cos θ {\displaystyle T_{s}(t)=cH\cos \theta }

missä H on sovellettu magneettinen vääntökenttä, θ {\displaystyle {\theta }}

on helmiäisen magneettisen momentin kulma alkuperäiseen magnetoitumissuuntaan nähden ja c on helmiäisvakio, joka löytyy kokeista, jotka on tehty sijoittamalla helmiäinen nesteeseen, jonka viskositeetti on tunnettu, ja soveltamalla siihen vääntökenttää.
Kompleksista dynaamista moduulia G voidaan käyttää esittämään värähtelevän jännityksen ja venymän välisiä suhteita:
G = G ′ + i G ″ {\displaystyle G=G’+iG”}
jossa G ′ {\displaystyle G’}

on varastointimoduuli ja G ″ {\displaystyle G”}

on häviömoduuli: G ′ = σ 0 ε 0 cos ϕ {\displaystyle G’={\frac {\sigma _{0}}{\varepsilon _{0}}}}\cos \phi }\.

G ″ = σ 0 ε 0 sin ϕ {\displaystyle G”={\frac {\sigma _{0}}{\varepsilon _{0}}}\sin \phi }

missä σ 0 {\displaystyle \sigma _{0}}

ja ε 0 {\displaystyle \varepsilon _{0}}

ovat jännityksen ja venymän amplitudit ja ϕ {\displaystyle {\displaystyle \phi }
on niiden välinen vaihesiirtymä.

Yllä olevista suhteista kompleksimoduulin komponentit määritetään silmukasta, joka syntyy vertaamalla vääntömomentin muutosta ajan muutokseen, joka muodostaa silmukan, kun sitä esitetään graafisesti. T s ( t ) {\displaystyle T_{s}(t)} raja-arvot.
– d(t) silmukan ja T s ( t ) {\displaystyle T_{s}(t)} rajoittaman alueen, A, rajoissa.
– d(t) silmukka, joka edustaa energiahäviötä sykliä kohti, käytetään laskelmissa. Vaihekulma ϕ \displaystyle \phi }
, varastointimoduuli G’ ja häviömoduuli G ovat tällöin: ϕ = sin – 1 4 A π Δ T s Δ d {\displaystyle \phi =\sin ^{-1}{\frac {4A}{\pi \Delta T_{s}\Delta d}}}

G ′ = Δ T s Δ d cos ϕ {\displaystyle G’={\frac {\Delta T_{s}}{\Delta d}}\cos \phi }

G ″ = Δ T s Δ d sin ϕ = 4 A π ω Δ d 2 {\displaystyle G”={\frac {\Delta T_{s}}{\Delta d}}\sin \phi ={\frac {4A}{\pi \omega \Delta d^{2}}}}

jossa d on siirtymä.
Kuvassa 3 esitetty hystereesi edustaa punasoluissa esiintyvää viskoelastisuutta. On epäselvää, liittyykö tämä kalvomolekyylien vaihteluihin vai solunsisäisten ATP-pitoisuuksien ohjaamaan metaboliseen toimintaan. Lisätutkimuksia tarvitaan näiden vuorovaikutusten täydelliseksi selvittämiseksi ja punasolujen taustalla olevien viskoelastisten muodonmuutosominaisuuksien valaisemiseksi.
Verisuonten vaikutuksetMuutos
Kun tarkastellaan veren viskoelastista käyttäytymistä in vivo, on otettava huomioon myös valtimoiden, kapillaarien ja laskimoiden vaikutukset. Veren viskositeetti vaikuttaa virtaukseen ensisijaisesti suuremmissa valtimoissa, kun taas elastisuus, joka piilee punasolujen elastisessa muodonmuutoskyvyssä, vaikuttaa ensisijaisesti valtimoissa ja kapillaareissa. Aaltojen etenemisen ymmärtäminen valtimoiden seinämissä, paikallisen hemodynamiikan ja seinämän leikkausjännitysgradientin ymmärtäminen on tärkeää sydän- ja verisuonitoiminnan mekanismien ymmärtämiseksi. Valtimoiden seinämät ovat anisotrooppisia ja heterogeenisiä, ja ne koostuvat kerroksista, joilla on erilaiset biomekaaniset ominaisuudet, mikä tekee niiden mekaanisten vaikutusten ymmärtämisen, joita valtimot tuovat veren virtaukseen, hyvin vaikeaksi.
Lääketieteelliset syyt paremmalle ymmärrykselleEdit
Lääketieteellisestä näkökulmasta verenkierron viskoelastisten ominaisuuksien tutkimisen tärkeys käy ilmeiseksi. Kardiovaskulaaristen proteesilaitteiden, kuten sydänläppien ja veripumppujen, kehittämisen myötä tarvitaan pulsoivan veren virtauksen ymmärtämistä monimutkaisissa geometrioissa. Muutamia konkreettisia esimerkkejä ovat veren viskoelastisuuden vaikutukset ja sen vaikutukset sykkivien veripumppujen testaamiseen. Veren viskoelastisuuden ja alueellisen ja globaalin aivoverenkierron välillä on dokumentoitu vahvoja korrelaatioita kardiopulmonaalisen ohitusleikkauksen aikana.
Tämä on myös viitoittanut tietä verianalogin kehittämiselle proteesilaitteiden tutkimiseksi ja testaamiseksi. Klassinen glyseriinin ja veden analogi antaa hyvän kuvan viskositeetista ja inertiavaikutuksista, mutta siitä puuttuvat todellisen veren elastiset ominaisuudet. Yksi tällainen verianalogi on ksantaanikumin ja glyseriinin vesiliuos, joka on kehitetty vastaamaan veren kompleksisen viskositeetin sekä viskoosisia että elastisia komponentteja.
Normaalit punasolut ovat muodonmuutoskykyisiä, mutta monet tilat, kuten sirppisolusairaus, vähentävät niiden kimmoisuutta, mikä tekee niistä vähemmän muodonmuutoskykyisiä. Punasoluilla, joiden muodonmuutoskyky on heikentynyt, on lisääntyvä virtauseste, mikä johtaa punasolujen aggregaation lisääntymiseen ja happisaturaation alenemiseen, mikä voi johtaa uusiin komplikaatioihin. Muodonmuutoskyvyltään heikentyneiden solujen läsnäolo, kuten sirppisolutaudissa, pyrkii estämään plasmakerrosten muodostumista, ja viskoelastisuutta mittaamalla eston aste voidaan määrittää määrällisesti.
Historia Muokkaa
Varhaisessa teoreettisessa työssä verta käsiteltiin ei-newtonilaisena viskoosina nesteenä. Ensimmäisissä tutkimuksissa oli arvioitu verta tasaisen virtauksen aikana ja myöhemmin värähtelevän virtauksen avulla. Professori George B. Thurston Texasin yliopistosta esitti ensimmäisen kerran ajatuksen veren viskoelastisuudesta vuonna 1972. Aiemmat tutkimukset, joissa tarkasteltiin verta tasaisessa virtauksessa, osoittivat, että veren elastiset ominaisuudet olivat merkityksettömän vähäiset, koska elastinen järjestelmä on varastoitunut vereen virtauksen alkaessa, joten sen läsnäolo jää piiloon, kun virtaus saavuttaa tasaisen tilan. Varhaisissa tutkimuksissa käytettiin tasaisessa virtauksessa havaittuja ominaisuuksia johtamaan ominaisuuksia epävakaisiin virtaustilanteisiin. Lääketieteellisten menetelmien ja laitteiden kehittyminen edellytti parempaa ymmärrystä veren mekaanisista ominaisuuksista.