Replikation
Kopiering: Cool för DNA, Cool för oss
När några av Shmoops klasskamrater gick i skolan kom deras lärare alltid på dem med att kopiera. Men klasskamraterna sa till sina lärare att det låg i deras DNA att kopiera. Hon kunde inte argumentera mot den logiken, så de hamnade aldrig i trubbel. Vilken trevlig liten historia.
Vilken parallell försöker vi dra med denna inte så subtila och hesa berättelse? DNA kopierar hela tiden, och det måste det göra om det vill fortsätta att skapa alla dessa celler. Och, åh, hur det gör det! DNA-replikation är kopiering av DNA så att replikerande celler har tillräckligt med DNA för dotterceller, eller de nya celler som härstammar från den ursprungliga cellen. Celldelning, eller att göra en cell till två nya celler, sker genom mitos för icke-sexuella celler, eller de celler som inte är involverade i… sex, och meios för sexuella celler, eller de celler som blir smutsiga snabbare än Vinny och Pauly D en bra kväll på Karma.
Det är bara en fas: Mitos eller meios
Trots att cellerna finns i alla former och storlekar, som en Benetton-reklam, finns det i princip två typer av celler:
- Könsceller (nej, inte de där sakerna i Amsterdam)
- Somatiska celler (icke-könsceller, som inte heller är de där sakerna i Amsterdam)
Könsceller är de celler som producerar avkomma hos eukaryoter som förökar sig sexuellt, och de delas in i följande:
- Spermceller som innehåller den manliga halvan av DNA
- Äggceller som innehåller den kvinnliga halvan av DNA

Det finns fyra huvudstadier i livscykeln för de flesta eukaryota celler (exklusive könsceller).
Dessa kallas
- Gap 1 (eller G1) fas
- Syntes (eller ”S”) fas
- Gap 2 (G2) fas
- Mitos
Och nej, The Gap sponsrar inte mitos. G1-, S- och G2-faserna kallas normalt för ”interfas”, främst av personer som studerar mitos. Fast om man får dem ensamma kallar de det för ”tråkig fas.”
G1
G1 är fasen i den eukaryota livscykeln efter mitos och före DNA-syntesen, som är S-fasen. Under mitosen har det mesta av den aktivitet som inte är mitos stängts av, och G1 är den period då allting börjar igen. G1 är som en galen sömnlös som väcker cellen genom att säga: ”Hej, jag har skurit av kuponger hela natten, dags att vakna och leka med mig!”
S-fas
S-fas är livscykelns DNA-replikeringsfas. Vi kommer att prata mer om detta senare, men om du inte kan vänta kan du klicka vidare. I den här fasen fördubblar cellen mängden DNA för att göra tillräckligt med DNA till dottercellerna, och cellen går från 2n till 4n, där n är antalet uppsättningar av kromosomer, eller ploidi. De flesta somatiska celler är 2n, eller diploida, medan könsceller är haploida, vilket innebär att de har hälften så mycket DNA som somatiska celler (vanliga celler).
G2 Fas
G2 är cellcykelns Jan Brady. Alla pratar alltid om ”Mitos, Mitos, Mitos!” och ingen bryr sig om stackars G2. Det är den period i cellcykeln där mer proteinsyntes sker och mikrotubuli tillverkas, vilket är viktigt för mitosen. ”Är det allt?” Frågar du? Japp. Egentligen inte så viktigt, vilket är anledningen till att många celler hoppar över detta steg. Grodceller hoppar över det, liksom många cancerceller. Stackars G2…
Mitos
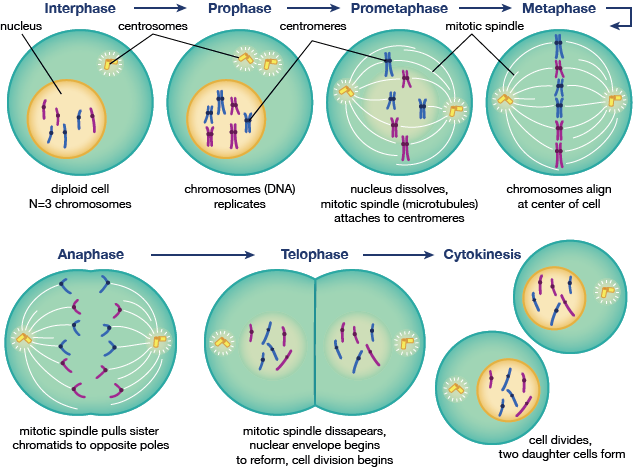
Mitos är det steg i cellcykeln som är så viktigt att det har en egen cykel. Mitos börjar efter G2 och börjar med profas, eller kondensering av kromatin till kromosomer och centrosomer som rör sig mot motsatta poler i cellen. Detta steg följs av prometafasstadiet, ett mellanstadium mellan profas och metafas, där kärnhöljet bryts ner och kromosomerna börjar samlas på mikrotubbar som förbinder centrosomerna. Metafas är det skede där alla kromosomer är uppradade i centrum, vilket följs av anafas, där kromosomerna delas på mitten och dotterkromatiderna flyttar sig till den närmaste centrosompolen. Telofas och cytokinesis är de sista stegen i celldelningen och återmontering av kärnan i två dotterceller, respektive.
Meoisis

Meiosen är M-fasen för könsceller. Den börjar som mitos och efter produktionen av de två första dottercellerna fortsätter den en andra mitoscykel. De 2n dottercellerna delar sig vidare för att producera 4 dotterceller, och en massa överprisade bröllop, var och en med n kromosomer.
I början…DNA-replikeringens ursprung
Om DNA aldrig replikerades skulle meios och mitos sakta halvera arvsmassans storlek tills varje cell skulle dö, vilket förmodligen inte skulle ta lång tid. Därför är det viktigt att DNA fördubblar sig självt för att ta hänsyn till att cellerna delar sig under mitos/meios. DNA-replikation liknar RNA-transkriptionen.
Nedan finns en tabell som jämför de två:
| DNA Replikation | RNA Transkription | ||
| Produkt | Dubbel-sträng DNA | mRNA | |
| Enzym | DNA-polymeras | RNA-polymeras | |
| Nukleotider Tillagd | Deoxynukleotidtrifosfater | Nukleotidtrifosfater | |
| Mallsträng | Botten | Botten, men transkriberar endast från antisense-strängen |
Vad kallar du de platser där replikationen börjar? Ursprung. Vi önskar att de hade kallat den Star Wars Episode I: The Phantom Menace, men det namnet var redan upptaget.
Originerna skiljer sig åt beroende på vilken typ av DNA det rör sig om. De flesta eukaryota celler och vissa bakterier och virus har linjärt DNA, vilket innebär att det finns en början och ett slut på DNA-sekvensen. Replikationens origins ligger därför i 5′-ändan av det linjära DNA:t. De flesta bakterier och vissa virus har cirkulärt DNA, vilket innebär att det finns en specifik sekvens som DNA-replikationsproteiner binder till (vilket i viss mån är analogt med RNA-transkription), och replikationen börjar från denna plats.

DNA-replikation börjar med att ett initiatorprotein binder, till exempel dnaA i E. coli-bakterier, eller ett komplex av proteiner, till exempel ursprungskomplexet i jäst. Initiatorernas funktion är att bryta isär de två DNA-strängarna så att replikationen kan börja. Denna situation är inte olik en domkraft som lyfter upp bilen när du byter däck. (Om du inte vet hur man byter däck kan du ringa en bärgningsfirma och fråga hur de gör. När de har berättat hur, skrik: ”Lägg på luren och be sedan att du aldrig behöver byta däck igen.
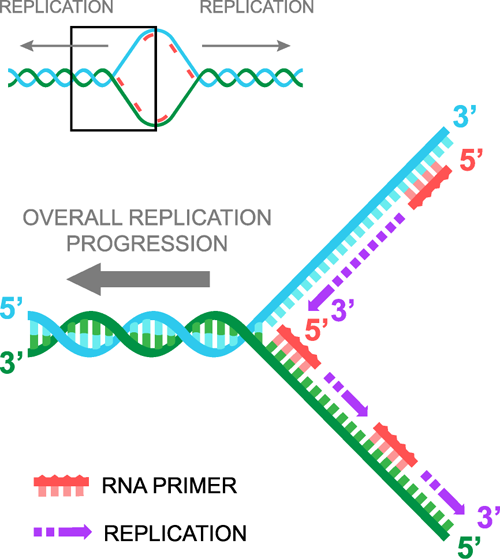
När DNA:t har skiljts åt kommer RNA-primers in, en binder sig till varje DNA-sträng och sedan fortsätter DNA-replikationen i en 5′- till 3′-riktning. Det är viktigt att inse att det är två 5′ till 3′-replikationer som sker samtidigt. Dessa två replikationer kallas var och en för en replikationsgaffel. Inte att förväxla med duplikationsskeden.

Passera replikationsgaffeln
När man delar DNA har vi en sträng som är 5′ till 3′ när vi tittar från vänster till höger, vilket vi kallar den ”eftersläpande” strängen, och en komplementär sträng som är 3′ till 5′ när vi tittar från vänster till höger, vilket vi kallar den ”ledande” strängen. När vi lägger till RNA-primers binder den ena primern till den ledande strängen och den andra primern till den efterföljande strängen. Den primer som binder till den ledande strängen är identisk med sekvensen i den efterföljande strängen, medan den primer som binder till den efterföljande strängen är identisk med sekvensen i den ledande strängen. Vi hoppas att ditt huvud inte har exploderat än.
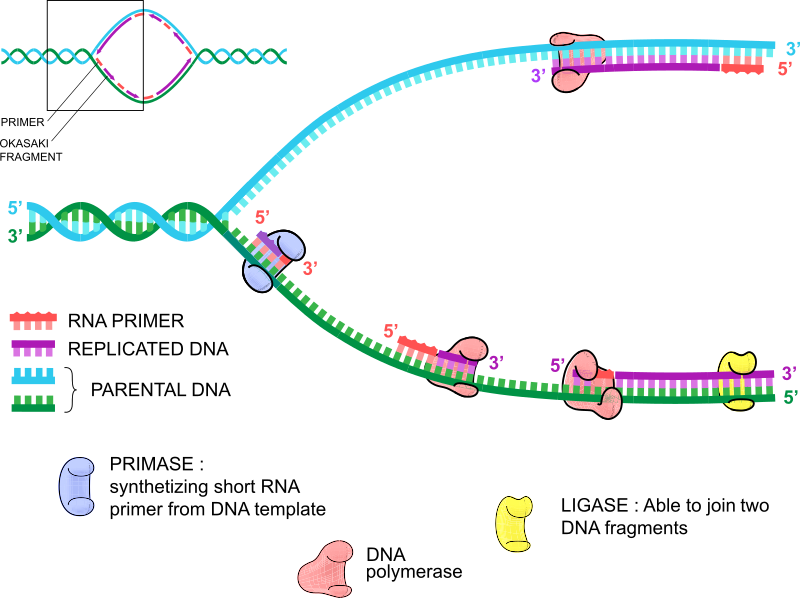
Tänk på DNA-replikation som din vänstra och högra hand. De är i princip likadana men har subtila skillnader. Din vänstra hand har tummen som femte finger om du räknar från vänster till höger, medan din högra hand har tummen som första finger. Ser du? Ledsen om vi blåste dig på näsan där. Precis som dina händer replikerar därför båda DNA-strängarna på samma sätt, men det finns en subtil skillnad. DNA-replikation fungerar endast i en 5′- till 3′-riktning, så två DNA-polymerasmolekyler binder RNA-primerna och börjar replikera i motsatt riktning. Replikationen av den ledande strängen driver replikationsprocessen eftersom RNA-primern driver replikationen av 5′- till 3′-strängen.
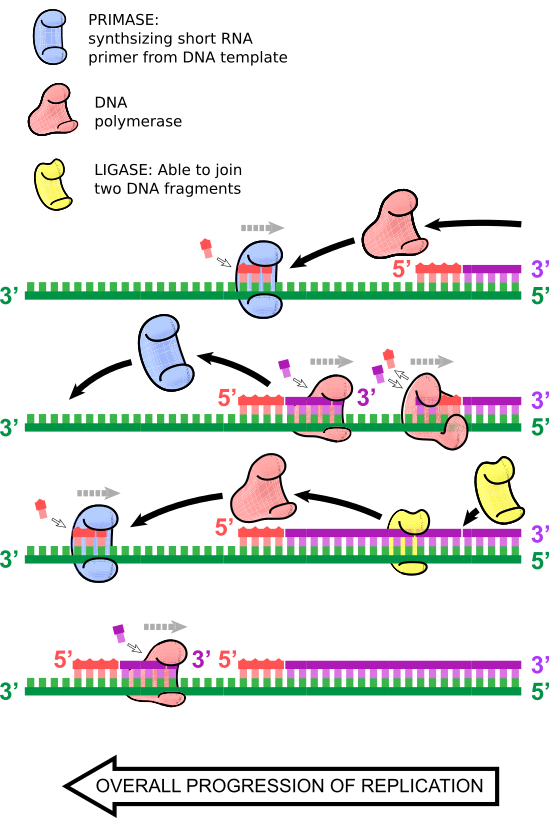
”Men Shmoop, hur replikerar man den ’eftersläpande’ strängen?”, kanske du frågar.
Som namnet antyder fortsätter DNA-replikationen av den ledande strängen under en viss tid tills en ny RNA-primer infogas, och ett andra DNA-polymeras startar från denna andra eftersläpande RNA-primer och replikerar DNA tills det träffar den första RNA-primern, eller den tidigare sträckan av dubbelsträngat DNA. Där faller DNA-polymeraset av, och DNA-ligaset förenar dessa DNA-fragment. Denna process fortsätter samtidigt med syntesen av den ledande strängen, så det finns många små replikerande dubbelsträngade DNA-fragment på den eftersläpande strängen. Dessa kallas Okazakifragment, efter makarna Reiji och Tsuneko Okazaki som upptäckte dem.
Är vi klara än?
DNA-replikationen slutar i bakterier och virus med cirkulära genomer när de två replikationsgafflarna möts på motsatt sida av kromosomen från replikationsursprunget, i ett magnifikt spel om kyckling. Det modiga replikationsterminatorproteinet går in och pausar DNA-replikationen så att de inte krockar med varandra. Termineringsmekanismen leder dock till att två sammanlänkade cirkulära kromosomer sitter fast i varandra som trollkarlsringar. Topoisomerasenzymer, såsom DNA-gyrase och topoisomeras IV, som ansvarar för att knäcka DNA, skär tillfälligt av den ena kromosomen så att den andra kan passera igenom. Där har vi det, vi förstör trolleritrick för alla.
Animation av linjär kromosomförkortning
Linjära kromosomer har en annan uppsättning problem eftersom vi med dessa killar aldrig når fullständig replikation av hela kromosomen. Varje replikationssteg förkortar DNA:t lite mer. Bakterier och virus med linjära kromosomer kringgår detta problem genom att antingen
- Initiera replikationen i mitten av kromosomen, eller genom att
- Primera replikationen med ett protein i stället för en RNA-primersekvens.
Eukaryoter kan dock inte göra någon av dessa och har utvecklat terminatorsekvenser som kallas telomerer som kringgår problemet med DNA-förkortning.

Endarna på linjärt eukaryotiskt DNA kallas telomerer, och även om de regelbundet förkortas varje replikationscykel, löser enzymer som kallas telomeraser problemet med DNA-förkortning. Telomererna är mycket repetitiva och telomeraserna lägger till korta sekvenser för att förlänga DNA-änden så att de inte blir för korta och så att viktiga delar av DNA-sekvensen inte går förlorade. Telomeraser är omvända transkriptaser, eller enzymer (vi kan se det på -ase) som omvandlar RNA till DNA. Se avsnittet ”Spiderman och andra exempel på rekombinant DNA” för mer information.
Telomeraser tar en kort RNA-sekvens, omvandlar den till dubbelsträngad DNA-sekvens och fäster den i slutet av DNA. Det finns en minimilängd som telomererna ska upprätthålla, och om DNA-sekvensen är kortare än minimilängden, kallad Hayflick-gränsen, dör cellen. Telomeraserna ser till att cellen kan fortsätta att replikera och dela sig i all oändlighet. Och överaktivitet av telomeraser är ofta kopplad till cancer, eftersom cellerna blir odödliga, som The Highlander.
Whoops, Mistakes Were Made: Du tänker förmodligen: ”DNA-polymeras tycker att det är så fantastiskt eftersom det replikerar allt DNA i min kropp så att jag kan överleva och fortsätta att genomgå mitos och meios.” Okej, vi är säkra på att de flesta av er inte tänker så, men låt oss spela med för den handfull människor som tänker så.
DNA-polymeras är faktiskt inte så fantastiskt! Det gör misstag precis som du eller Shmoop – även om våra misstag oftast är produkter av för många femtimmars energidrycker blandade med friterade Twinkies. DNA-polymeraset måste dock rätta till dessa misstag, medan vi på Shmoop går in i en sockerinducerad koma.

I brådskan med DNA-replikationen läggs ibland en felaktig bas till sekvensen, som inte basparar sig korrekt med mallsträngen. Därför får vi konstiga G-T- eller A-G-baspar. Vi kommer alla ihåg att normala par är G-C och A-T (Eh? Vad är det?). Dessa felaktiga par är inte bra för oss eftersom det innebär att våra gener muteras och vi blir mutanter. Få mutationer leder någonsin till häftiga saker som teleportering eller lasersyn… eller något från X-Men. Shucks.

Stora eukaryoter som vi som replikerar långsamt och lever länge vill inte att mycket mutationer ska ske. Varför inte? Alla mutationer som påverkar vår tillväxt är vanligtvis … inte bra … och kommer att skada oss. Sorgligt ansikte.
Mindre organismer, som bakterier, virus och till och med flugor, har dock korta livscykler i storleksordningen dagar och ibland timmar. De replikerar sig som galningar, där varje ny generation kommer att ha 100-tals-1000-tals fler individer än den föregående generationen. Därför kan mutationer i deras fall vara mer fördelaktiga. Ibland vill de undkomma bekämpningsmedel, antibiotika eller läkemedel som utformats för att behandla/döda dem, vilket är anledningen till att det är så svårt att tillverka ett läkemedel som hindrar oss från att bli sjuka. Bakterier och virus fortsätter att mutera och replikera. Och som vi redan har märkt finns det lite felkontroll i deras polymeraser.
Hjärnsnack
Här är en snygg video om hur DNA-replikation faktiskt fungerar.
Kan du upptäcka DNA-polymeraserna? De är de flerfärgade sfärerna, en för den ledande strängen och en för den efterföljande strängen. De eftersläpande strängpolymeraserna är de som kommer in och sedan faller av när dubbelsträngen är färdigställd.