14.1 Inleiding tot neuropeptiden en stikstofmonoxide
Klassieke neurotransmitters (b.v. glutamaat en ACh) werken door de gerichte overdracht van informatie tussen presynaptische releaseplaatsen en postsynaptische clusters van receptoren. Het proces is over het algemeen ruimtelijk goed georganiseerd met enige, maar zeer beperkte, “spill-over” van de zender tussen naburige synapsen. De situatie is heel anders voor transmissie die door neuropeptiden en stikstofmonoxide (NO) wordt bemiddeld. Deze moleculen negeren de richting van de informatiestroom en beïnvloeden in plaats daarvan een volume van het gebied vanaf het punt waar zij vrijkomen. Dit kenmerk is losjes analoog aan het vrijkomen van peptidehormonen in de bloedstroom, die naar verafgelegen plaatsen kunnen worden gebracht om daar hun effect te sorteren. Zo kan NO worden geproduceerd in een postsynaptisch neuron en terugdiffunderen naar het presynaptische neuron of naar andere naburige neuronen, waar het de daaropvolgende afgifte van een transmitter beïnvloedt. Deze eigenschap wordt retrograde transmissie genoemd, omdat het signaal in de tegenovergestelde richting reist (post- naar pre-) in vergelijking met de klassieke richtinggevoeligheid van neurotransmissie.
14.2 Neuropeptiden
Veel neuropeptiden werden oorspronkelijk beschreven als hormonen (b.v. somatostatine en cholecystokinine), hoewel zij, eenmaal gekarakteriseerd, vervolgens ook werden aangetroffen in neuronen binnen het CZS. Neuropeptiden veroorzaken neuronale reacties met een langzame aanvang en een lange duur (Figuur 14.1), en elke gekarakteriseerde neuropeptide-receptor valt in de GPCR-klasse (G-eiwit gekoppelde receptor). Ter vergelijking toont figuur 14.1 typische reacties die worden opgewekt door de werking van een klassieke neurotransmitter (glutamaat) die werkt op een ionotrope receptor en een neuropeptide. Een actiepotentiaal opgewekt in neuron 1 (in groen), een glutamaat-afgevend neuron, produceert de snelle (5 msec) EPSP die typisch is voor glutamaat-gemedieerde synaptische transmissie. Daarentegen produceert een actiepotentiaal opgewekt in neuron 2 (paars), dat neuropeptide vrijgeeft, een traag begin (5 sec tot piek) en lange duur (10 sec) EPSP. Deze langzame EPSP’s zijn typerend voor neuropeptidehandelingen, aangezien alle neuropeptidereceptoren aan G-eiwitten gekoppeld zijn.
Figuur 14.1
Verder brengen veel neuropeptiden geen duidelijke elektrofysiologische verandering in het postsynaptische neuron teweeg. Bijvoorbeeld, een actiepotentiaal in neuron 3 (figuur 14.1; roze) geeft geen respons in het postsynaptische neuron, ook al kan men met andere middelen bevestigen dat er neuropeptiden zijn vrijgemaakt. Er hebben zich echter veranderingen voorgedaan in het postsynaptische neuron, zoals blijkt uit het voorbeeld in de panelen met het label “C”. Als men eerst de glutamaatrespons (C1) meet door neuron 1 te stimuleren, wordt een typische respons gezien. Stimulatie van neuron 3 geeft geen respons (C2) zoals voorheen. Indien echter neuron 1 opnieuw wordt gestimuleerd na stimulatie van neuron 3, wordt het EPSP gemeten in het postsynaptische neuron versterkt (C3) (het oorspronkelijke EPSP is de ononderbroken lijn en het EPSP na stimulatie van neuron 3 is de gestippelde lijn). Het neuropeptide dat vrijkwam uit neuron 3 veranderde dus de reactie van het postsynaptische neuron op een andere neurotransmitter. Van deze neuropeptiden wordt gezegd dat zij neuromodulatoren zijn, omdat zij de effecten van een tweede transmitter versterken of onderdrukken. Andere voorbeelden van dergelijke heterosynaptische plasticiteit worden beschreven in hoofdstuk 7, deel 2.
14.3 Indeling van peptiden naar families
Neuropeptiden kunnen worden gegroepeerd in families op grond van overeenkomsten in hun aminozuursequenties (Tabellen I en II).
Tabel I
Neuropeptidefamilies
Tachykininen: substance P, bombesine, stof
Insulinen: insuline, insuline-achtige groeifactoren
Somatostatinen: somatostatine, pancreas-polypeptide
Gastrinen: gastrine, cholecystokinine
Opioïden: opiocortinen, enkefalinen, dynorfine
Zo worden de opiaten als familie gegroepeerd wegens de identieke aminozuursequentie Tyr-Gly-Gly-Phe–(tabel II) die aan het N-uiteinde van elk wordt aangetroffen.
| Tabel II Opiatenfamilie – Identiteit van de sequentie in cursief |
|
| Naam | Aminozuursequentie |
| Leu-enkephalin | Tyr-Gly-Gly-Phe-Leu-OH |
| Met-enkephalin | Tyr-Gly-Gly-Phe-Met-OH |
| Beta-endorphin | Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys- Ser-Gln-Thr-Pro-Leu-Val-Thr-Leu- Phe-Lys-Asn-Ala-Ile-Val-Lys-Asn-Ala- His-Lys-Gly-Gln-His-OH |
| Dynorfine | Tyr-Gly-Gly-Phe-Leu-Arg-Ile-Arg- Pro-Lys-Leu-Lys-Trp-Asp-Asn-Gln-OH |
14.4 Biosynthese en regulering
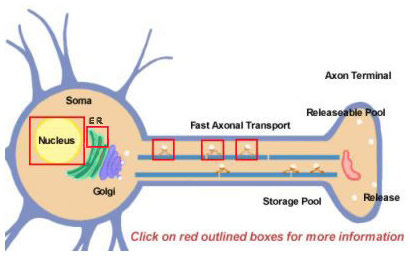
Figuur 14.2
Neuropeptiden worden afgeleid van grotere precursoren door proteolytische verwerking. Zij worden alle in eerste instantie gesynthetiseerd in het soma (figuur 14.2). Precursors worden aanvankelijk gevormd door translatie op polyribosomen die aan het endoplasmatisch reticulum nabij het cellichaam zijn bevestigd. Van daaruit passeren zij het Golgi-apparaat waar verdere modificaties plaatsvinden, waaronder glycosylering. Vervolgens worden ze verpakt in secreetkorrels die naar de uiteinden worden vervoerd via snel axonaal transport. Snel is echter een relatief begrip, gezien de relatief lange afstanden die sommige neuropeptiden moeten afleggen. Het aanvullen van de pool van vrij te geven neuropeptiden kan vele uren duren. Tijdens het transport naar het zenuwuiteinde beginnen proteasen, die in de blaasjes zijn verpakt, het neuropeptide te splitsen in zijn uiteindelijke rijpe vorm. Deze verwerking is essentieel voor de activering van de neuropeptiden, aangezien de precursoren biologisch inactief zijn. Ten minste drie soorten verwerking vinden plaats in de blaasjes (Figuur 14.2; klik op de kadertjes voor meer details). Eerst splitst een endopeptidase de precursor om twee nieuwe producten te genereren (Eerste kader in axon). Voor veel precursors vindt deze splitsing plaats na basische residuen (zoals Lys en Arg) en wordt uitgevoerd door trypsine-achtige proteasen. Vervolgens, hoewel niet voor elk neuropeptide, klieft een carboxypeptidase de basische residuen van de C-terminus van het nieuwe peptide (Tweede vakje in axon). Tenslotte zet een derde enzym de COOH-groep (carboxylgroep) van een Gly-residu, dat in de C-terminus van veel neuropeptiden voorkomt, om in een NH2-groep (amidegroep) om de rijpe, actieve vorm van het neuropeptide te produceren.
14.5 Meerdere mechanismen worden gebruikt om de diversiteit van neuropeptiden te produceren
De meeste eiwitten worden geproduceerd uit mRNA-moleculen die van precursor-RNA’s worden gesplitst in hun uiteindelijke vorm in de kern. Differentiële splicing is één manier die een neuron gebruikt om de productie van verschillende soorten neuropeptiden te diversifiëren. Een bekend voorbeeld is het mRNA voor stof P dat normaliter ook mRNA bevat dat codeert voor stof K. Het stof K-gedeelte van het mRNA kan differentieel worden gesplitst, zodat het resulterende mRNA alleen stof P kan produceren (klik op het kader boven de kern in figuur 14.2)
Figuur 14.3
Neuropeptiden worden geproduceerd uit een langer precursoreiwit door proteolytische verwerking. Een uitstekend voorbeeld is de opioïdenfamilie van peptiden (b.v. de verwerking van proopiomelanocorticotropine, POMC en Enkefaline; zie figuur 14.3). Zoals gezegd, vindt de proteolytische verwerking plaats binnen de transportblaasjes en meestal door splitsing van de precursor aan de N-terminale zijde van basische residuen (arginine en lysine), hoewel ook andere splitsingsplaatsen zijn geïdentificeerd. In sommige gevallen, zoals bij het Enkefaline-precursoreiwit, zijn er meerdere kopieën van hetzelfde uiteindelijke bioactieve peptide aanwezig. De ene precursormolecule bovenaan in figuur 14.3 bevat zes kopieën van Met-enkefaline (ME) en één kopie van Leu-enkefaline (LE).
Diversiteit kan dus worden gegenereerd door de volgorde van de splitsingsplaatsen te veranderen door differentiële splitsing, door verschillende proteasen te produceren en/of te verpakken (die verschillende sequenties voor splitsing herkennen) in de transportblaasjes, of door een proteolytische plaats te verbergen door post-translationele modificaties. Een voorbeeld van dit laatste is dat een specifieke splitsingsplaats kan worden verborgen door toevoeging van een koolhydraatzijketen die de toegang van het protease tot die plaats sterisch blokkeert. Een andere veel voorkomende bevinding is dat één enkel precursormolecuul verschillende neuropeptiden zal bevatten (zie figuur 14.3) en dat derhalve de soorten bewerking die plaatsvinden uiteindelijk bepalen welk neuropeptide door het neuron wordt vrijgemaakt. Het POMC-precursoreiwit kan worden gesplitst tot ACTH (oranje) en β-lipoproteïne (lichtblauw), die elk verder kunnen worden gesplitst om aanvullende bioactieve neuropeptiden te genereren (figuur 14.3). Zo kan bijvoorbeeld het β-lipoproteïne (lichtblauw) verder worden gesplitst in zowel γ-lipoproteïne (groene strepen) als β-endorfine (donkerblauw). Nogmaals, afhankelijk van de verwerking die plaatsvindt, kan hetzelfde precursor-eiwit worden gewijzigd om neuropeptiden te produceren met dramatisch verschillende biologische reacties.
14.6 Vrijkomen
Figuur 14.4
Peptiden komen vrij door calcium-afhankelijke exocytose met enkele belangrijke verschillen met het vrijkomen van klassieke neurotransmitters. Typisch zijn de blaasjes die neuropeptiden vrijgeven veel groter dan die welke kleine molecule neurotransmitters bevatten (b.v. glutamaat) en vereisen geen presynaptische specialisatie voor vrijgave (zie de elektronenmicrografiek in figuur 14.4). In tegenstelling tot de kleine blaasjes die glutamaat bevatten, lijken de grote blaasjes niet gedockt aan het membraan. Deze waarneming is in overeenstemming met het idee dat kleine molecule neurotransmitters korte, plaatselijke effecten veroorzaken (bij synaptische verbindingen), terwijl neuropeptiden langzame, langdurige effecten veroorzaken die vaak een aanzienlijk gebied rond de plaats van afgifte omvatten. Bedenk ook dat, aangezien neuropeptiden in het soma van de cel worden gesynthetiseerd en niet plaatselijk bij de synaps, als hun voorraad uitgeput is door aanhoudende afgifte, het verscheidene uren kan duren om de vrij te geven pools weer aan te vullen. Bijvoorbeeld, een motorneuron, met zijn cellichaam in het ruggenmerg en de synaps in de voet, heeft een axon van een meter lang. Als gebruik wordt gemaakt van snel axonaal transport zou het meer dan een dag kunnen duren voordat een nieuw gesynthetiseerd neuropeptide vanuit het soma bij deze synaps aankomt. Het zou ook duidelijk moeten zijn dat endogene pijndodende neuropeptiden, zoals bèta-endorfine, “opgebruikt” zouden kunnen worden in tijden van aanhoudende stimulatie, leidend tot situaties waarin pijn niet langer kan worden beheerst door endogene mechanismen.
Een typisch volgroeid neuron zal vaak één kleine molecule neurotransmitter en één of meer neuropeptiden vrijgeven (zoals in het voorbeeld van figuur 14.4). Als er meer dan één neuropeptide vrijkomt, is dat meestal afkomstig van hetzelfde precursormolecuul. Een voorbeeld is de gezamenlijke vrijlating van zowel ACh als calcitonine-gen-gerelateerd peptide uit spinale motorneuronen. CGRP activeert adenylaat-cyclase, waardoor het cAMP-niveau stijgt, en versterkt de contractiekracht die wordt opgewekt door ACh-activering van de nicotinische ACh-receptor. In dit geval is het neuropeptide modulerend zoals beschreven in figuur 14.1. In dit geval versterkt het effect echter de spiercontractie in plaats van de grootte van de EPSP te vergroten. In beide voorbeelden is de versterkte respons het gevolg van een verhoogde gevoeligheid van het systeem voor een constante hoeveelheid vrijgekomen neurotransmitter.
14.6 Beëindiging van de werking
Neuropeptiden worden langzaam uit de extracellulaire ruimte verwijderd; een kenmerk dat ook bijdraagt tot hun relatief langdurige effecten. Inactivering gebeurt zowel door diffusie als door afbraak door extracellulaire proteasen. Er zijn geen aanwijzingen gevonden voor de heropname van peptiden als middel om hun werking te beëindigen.
14.7 Receptoren zijn alle gekoppeld aan G-eiwitten
Alle bekende neuropeptidereceptoren produceren hun effecten door de niveaus van intracellulaire tweede boodschappers te veranderen. Deze receptoren zijn zeven transmembraan-overspannende proteïnen die via G-proteïnen (GPCR’s) verbonden zijn om de activering van andere cellulaire enzymen te veranderen. Deze eigenschap komt overeen met neuropeptiden die een tragere reactie induceren en is zeer geschikt voor een modulerende rol. Een belangrijk onderscheid tussen kleine en neuropeptide moleculaire transmitters is dat neuropeptide receptoren een hoge affiniteit hebben voor binding (nanomolair) in tegenstelling tot micro- of millimolaire affiniteiten gemeten voor kleine moleculaire neurotransmitters (zoals glutamaat). Aangezien neuropeptiden niet gericht worden vrijgelaten in het begrensde volume van een synaps, bereiken hun concentraties geen zeer hoge niveaus en de receptoren moeten dan een hoge affiniteit hebben om op deze kleine concentraties te reageren. Deze hoge affiniteit vertraagt de dissociatie van het neuropeptide van zijn receptor en draagt ook bij tot de persistente effecten van deze moleculen.
14.8 Nitrisch Oxide (NO)
Nitrisch oxide heeft wijdverspreide aandacht gekregen als het stichtend lid van een nieuwe klasse van gasvormige boodschappermoleculen. NO is de actieve molecule die sublinguale nitroglycerine produceert om de vasodilatatie te verhogen bij de verlichting van angina pectoris. Andere belangrijke biologische effecten van NO worden nu erkend. In het zenuwstelsel is NO bijvoorbeeld belangrijk voor de regulering van de hersendoorbloeding, voor de modulatie van de neurotransmissie en voor de toxiciteit bij diverse pathologische toestanden.
14.9 Eigenschappen van NO
Samenvatting van de eigenschappen van NO
- Gas dat vrij door membranen diffundeert
- Kortlevend met een halveringstijd gemeten in seconden
- Kortlevend met een halveringstijd gemeten in seconden
- .levensduur gemeten in seconden
- Zeer reactieve vrije radicaal
- Giftig bij hoge concentraties
NO is een kortlevend gas dat niet moet worden verward met het relatief stabiele verdovingsgas lachgas. NO is eigenlijk een vrije radicaal en is dus een zeer reactieve verbinding. Sommige van zijn toxische effecten zijn waarschijnlijk te wijten aan de reactie van NO met superoxide, waarbij het destructieve radicaal peroxynitraat ontstaat. NO wordt beschouwd als een onconventionele neurotransmitter, omdat het niet wordt vrijgemaakt door exocytose en zijn werking niet plaatsvindt via conventionele receptormoleculen.
Zoals eerder vermeld, wordt er bij de typische beschrijving van neuronale communicatie van uitgegaan dat de transmissie in één richting verloopt. Er komt een presynaptische neurotransmitter vrij die veranderingen teweegbrengt in het postsynaptische neuron. Verscheidene verbindingen (zoals neuropeptiden en NO) die in postsynaptische neuronen worden geproduceerd, diffunderen naar de lokale omgeving en beïnvloeden de omringende cellen. Aangezien NO een vrij verspreidbaar gas is, kan het zich vanaf het punt van productie snel in alle richtingen verplaatsen. Als bijvoorbeeld door stimulatie met een glutamaatreceptor NO wordt geproduceerd in een postsynaptische cel, kan het vrijkomen in de lokale omgeving en een signaal terugzenden naar het presynaptische neuron (figuur 14.5). Dit type activiteit wordt retrograde signalering genoemd, omdat het signaal in retrograde richting van de postsynaptische naar de presynaptische neuron gaat.
Figuur 14.5
Figuur 14.6
De figuren 14.5, 14.6 en 14.7 geven een overzicht van de belangrijkste aspecten van de NO-synthese. In dit voorbeeld komt glutamaat vrij uit de presynaptische terminal, dat zich bindt aan NMDA-receptoren op het postsynaptische membraan, waardoor deze opengaan en een Ca2+-influx mogelijk wordt. Het Ca2+ activeert calmoduline dat zich bindt aan het enzym stikstofoxidesynthase (NOS) en dit activeert. Met arginine als substraat produceert NOS NO en een tweede reactieproduct citrulline. Het NO is dan vrij om zich in de omgeving te verspreiden en een wisselwerking aan te gaan met de presynaptische terminal die het glutamaat aanvankelijk heeft vrijgemaakt of met een andere cel in de lokale omgeving. Besef dat elk proces dat intracellulair Ca2+ verhoogt, mogelijk de NOS zal activeren. Glutamaat activatie van NMDA receptoren is slechts één goed gedocumenteerd voorbeeld.
Figuur 14.7
14.10 Synthese door stikstofoxidesynthase (NOS) en afgifte
NO wordt geproduceerd door het enzym stikstofoxidesynthase (NOS). Dit enzym wordt aangetroffen in een subpopulatie van neuronen (1-2% van de neuronen in cortex) en wordt aangetroffen in de meeste endotheelcellen. Ten minste één vorm van NOS in deze cellen is afhankelijk van calcium en calmoduline voor activering, zoals aangegeven in de figuren 14.5, 14.6 en 14.7. Zij bevat ook cofactoren die lijken op die van cytochroom P-450. Deze cofactoren zijn NADPH (nicotinamide adenine dinucleotide fosfaat, FAD (flavin adenine mononucleotide) en FMN (flavin mononucleotide). Deze cofactoren zijn essentieel voor de overdracht van elektronen die het onstabiele en kortlevende product NO voortbrengt. Het substraat in deze reactie is het gewone aminozuur arginine en de producten zijn citrulline en NO (figuur 14.5). Door de korte levensduur van NO is het buitengewoon moeilijk rechtstreeks te meten. In experimentele preparaten is er echter een uitstekende correlatie tussen de toediening van NMDA, dat intraneuronaal Ca2+ verhoogt en NOS activeert, en de productie van het bijkomende product van de enzymatische activiteit van NOS, citrulline (zie figuur 14.8). De citrullineproductie is een betrouwbare indicator van de NO-productie. Het is ook mogelijk de productie van citrulline (en NO) te blokkeren door cellen het niet-metaboliseerbare substraat voor NOS, methylarginine genaamd, te voeren (figuur 14.9).9), en dergelijke verbindingen zijn gebruikt om de productie van NO te verminderen en de biologische effecten ervan te beëindigen.
Figuur 14.8
Figuur 14.9
14.11 “Receptoren” voor NO
Figuur 14.10
Een van de belangrijkste doelwitten voor NO blijkt het enzym guanylyl cyclase te zijn. Deze ontdekking werd indirect gedaan door de accumulatie van het NOS-product citrulline te controleren en tegelijkertijd te letten op de productie van cGMP, het product van het enzym guanylyl cyclase (figuren 14.8 en 14.9). In dit experiment werd NMDA toegevoegd aan een neuronaal preparaat om de NOS te activeren. NOS gebruikt arginine als substraat om NO te produceren en zoals gezegd, indien een niet-hydrolyseerbaar analoog zoals methyl-arginine aan het systeem wordt toegevoegd, bleken citrulline en cGMP-accumulatie op dezelfde dosis-afhankelijke wijze te eindigen. Deze sterke correlaties leidden tot de ontdekking dat guanylyl cyclase een belangrijk NO-doelwit is.
Guanylyl cyclase is een ongewoon enzym, omdat het een heemring met een geassocieerd ijzermolecuul als onderdeel van zijn structuur heeft. NO activeert dit enzym door zich aan het ijzer in het heem te binden, waardoor de productie van cyclisch GMP uit GTP op gang komt (figuur 14.10) via conformatieveranderingen in het enzym. cGMP activeert vervolgens een cGMP-afhankelijk proteïnekinase en andere enzymen die hieronder worden beschreven. Het spectrum van effecten die door de activering van cGMP-afhankelijke proteïnekinase worden teweeggebracht, wordt nu pas goed begrepen.
14.12 Biologische effecten van NO
NO veroorzaakt een grote verscheidenheid aan biologische effecten. Helaas zijn wij bijzonder onwetend over de rol van NO bij de modulatie van cellulaire processen in het zenuwstelsel. De rol van NO bij de regulering van de bloedvaten is goed gedocumenteerd en het lijkt een soortgelijke belangrijke rol te spelen in het zenuwstelsel.
14.13 Vasodilatator
Figuur 14.11
Klik hier om een vergrote afbeelding van deze afbeelding te zien.Onder normale omstandigheden draagt NO bij tot de regeling van de bloedstroom door het cerebrovasculaire systeem. Er bestaat noodzakelijkerwijs een snel terugkoppelingsmechanisme om actievere delen van de hersenen van de nodige voedingsstoffen te voorzien. Dit mechanisme is noodzakelijk omdat de hersenen een zwakke reserve aan energievoorraden hebben. NO wordt geproduceerd in neuronen die NOS bevatten en een aanhoudende activiteit ondergaan. Deze omstandigheden bevorderen de activering van NMDA-receptoren, waarvan bekend is dat zij NO-productie veroorzaken. NO diffundeert van deze gelokaliseerde gebieden met hoge neuronale activiteit naar de omringende microvasculatuur (figuur 14.11), waardoor vaatverwijding en een verhoogde bloedstroom ontstaan.
Hoewel de exacte mechanismen waarmee NO vaatverwijding teweegbrengt nog niet zijn gedefinieerd, is bekend dat activering van cGMP-afhankelijke proteïnekinase in gladde spiercellen een ontspanning van de vaten teweegbrengt. Aangezien een van de belangrijkste doelwitten van NO guanylylcyclase is (dat cGMP produceert en cGMP-afhankelijk proteïnekinase activeert), wordt aangenomen dat een belangrijke route voor de vaatverwijdende werking van NO via cGMP-afhankelijk proteïnekinase verloopt. Activering van dit kinase leidt indirect tot verlaagde Ca2+-niveaus in de gladde spiercellen en vervolgens tot defosforylering van het myosine-contractielapparaat, waardoor relaxatie optreedt (figuur 14.12). In gladde spiercellen blijkt NO de cellen ook rechtstreeks te hyperpolariseren, mogelijk door activering van K+-kanalen, hetgeen leidt tot de secundaire sluiting van Ca2+-kanalen, die eveneens spierrelaxatie teweegbrengt. Kortom, een van de belangrijkste functies van NO lijkt de integratie te zijn van het niveau van de neuronale activiteit met plaatselijke veranderingen in de cerebrale bloedstroom om een adequate perfusie van metabolisch actief weefsel te handhaven.
Figuur 14.12
14.14 Neuromodulator
Van NO wordt ook gedacht dat het werkt als een plaatselijk diffusibele boodschapper. Het wordt geproduceerd door elke actie die Ca2+ verhoogt in cellen die NOS bevatten, zoals glutamaatstimulatie van NMDA-receptoren. Via de daaropvolgende activering van guanylyl cyclase en de productie van cGMP beïnvloedt de NO-productie een reeks secundaire processen. Daartoe behoren directe modulatie van ionenkanalen, stimulatie van cGMP-afhankelijk proteïnekinase, en zowel up- als down-regulatie van cAMP-fosfodiësterase. Stroomafwaartse effecten zijn vervolgens talrijk en omvatten opwaartse en neerwaartse regulatie van Ca2+-kanalen, verhoogde prikkelbaarheid (verhoging van de neuronale vuursnelheid), verhoogde of verlaagde afgifte van neurotransmitters, en veranderingen in de neuronale morfologie.
14.15 Toxiciteit
NO in overmaat is giftig voor cellen. Er bestaat echter een paradox voor de toxiciteit van NO. Cellen die veel NO produceren zijn resistent tegen de toxische effecten ervan. Zo wordt de giftigheid van NO door macrofagen en neutrofielen gebruikt als een mechanisme om tumorcellen en bacteriën te doden. Geen van beide celtypes die NO produceren, is echter vatbaar voor de schadelijke effecten ervan. Deze bevinding geldt ook voor neuronen in het centrale zenuwstelsel. Een overmaat aan glutamaat leidt tot neurotoxiciteit in de hersenen en wordt beschouwd als de voornaamste oorzaak van neuronale sterfte bij ziekten als Huntington of Alzheimer of na een acute beroerte of trauma. Overmatige productie van NO wordt geacht een rol te spelen bij dit neuronale verlies, omdat het giftig is wanneer het in overmaat wordt geproduceerd. Interessant is dat cellen die positief kleuren voor het enzym NOS gespaard blijven in de degenererende delen van de hersenen die door deze ziekten zijn aangetast. De weerstand van deze cellen lijkt vergelijkbaar te zijn met de weerstand van de hierboven beschreven immuuncellen.
Test je kennis
- Vraag 1
- A
- B
- C
- D
- E
Neuropeptiden zijn aanwezig in synaptische terminals omdat ze:
A. Gesynthetiseerd in de terminal.
B. Vervoerd door snel axonaal transport
C. Vervoerd door diffusie
D. Vervoerd door langzaam axonaal transport
E. Opgenomen uit de extracellulaire ruimte
Neuropeptiden zijn aanwezig in synaptische terminals omdat ze:
A. Gesynthetiseerd in de terminal. Dit antwoord is INCORRECT.
De synthetische machinerie die nodig is voor de synthese van peptiden en eiwitten is niet te vinden in de axon terminal.B. Vervoerd door snel axonaal transport
C. Gedragen door diffusie
D. Vervoerd door langzaam axonaal transport
E. Opgenomen uit de extracellulaire ruimte
Neuropeptiden zijn aanwezig in synaptische terminals omdat ze:
A. Gesynthetiseerd in de terminal.
B. Vervoerd door snel axonaal transport Dit antwoord is CORRECT!
Neuropeptiden worden gesynthetiseerd in het cellichaam en worden verwerkt in blaasjes die langs microtubuli naar axonale terminals worden vervoerd via snel axonaal transport.C. Vervoerd door diffusie
D. Vervoerd door langzaam axonaal transport
E. Opgenomen uit de extracellulaire ruimte
Neuropeptiden zijn aanwezig in synaptische terminals omdat ze:
A. Gesynthetiseerd in de terminal.
B. Vervoerd door snel axonaal transport
C. Vervoerd door diffusie Dit antwoord is INCORRECT.
Diffusie is bovendien een uitstekend mechanisme om moleculen over korte afstanden (enkele microns) te verplaatsen, maar het is inefficiënt bij het verplaatsen van moleculen over afstanden die doorgaans het neuronale soma van de axonterminals scheiden.D. Vervoerd door langzaam axonaal transport
E. Opgenomen uit de extracellulaire ruimte
Neuropeptiden zijn aanwezig in synaptische terminals omdat ze:
A. Gesynthetiseerd in de terminal.
B. Vervoerd door snel axonaal transport
C. Vervoerd door diffusie
D. Getransporteerd door langzaam axonaal transport Dit antwoord is INCORRECT.
Ook is het belangrijk om de voorraad neuropeptiden aan de terminals zo snel mogelijk aan te vullen. In dit geval, sneller is beter.E. Opgenomen uit de extracellulaire ruimte
Neuropeptiden zijn aanwezig in synaptische terminals omdat ze:
A. Gesynthetiseerd in de terminal.
B. Vervoerd door snel axonaal transport
C. Vervoerd door diffusie
D. Vervoerd door langzaam axonaal transport
E. Opgenomen uit de extracellulaire ruimte Dit antwoord is INCORRECT.
Er is geen up-take mechanisme bekend om neuropeptiden uit de extracellulaire ruimte te verwijderen. Hun werkingsmechanisme wordt beëindigd door diffusie en door enzymatische afbraak door extracellulaire proteasen.>
- Vraag 2
- A
- B
- C
- D
- E
Nitric Oxide:
A. Heeft een lange levensduur
B. Is membraan doorlaatbaar
C. Is niet reactief
D. Is ook bekend als lachgas.
E. Is niet giftig voor cellen.
Nitric Oxide:
A. Heeft een lange levensduur Dit antwoord is INCORRECT.
NO heeft een halveringstijd gemeten in seconden en is zeer onstabiel.B. Is membraan doorlaatbaar.
C. Is niet-reactief.
D. Is ook bekend als lachgas.
E. Is niet giftig voor cellen.
Nitric Oxide:
A. Heeft een lange levensduur
B. Is membraan doorlaatbaar. Dit antwoord is CORRECT!
NO is vrij doorlaatbaar voor membranen en beïnvloedt alle cellen binnen een bepaalde straal vanaf het punt van productie.C. Is niet-reactief
D. Is ook bekend als lachgas.
E. Is niet giftig voor cellen.
Nitric Oxide:
A. Heeft een lange levensduur
B. Is membraan doorlaatbaar.
C. Is niet reactief Dit antwoord is INCORRECT.
NO is een zeer reactieve vrije radicaal. Het reageert met eiwitten, lipiden en nucleïnezuur en heeft een bijzonder hoge affiniteit voor gereduceerde metalen (zoals Fe2+ in de heemgroep van guanylyl cyclase).D. Is ook bekend als lachgas.
E. Is niet giftig voor cellen.
Nitric Oxide:
A. Heeft een lange levensduur
B. Is membraan doorlaatbaar
C. Is niet reactief
D. Is ook bekend als lachgas. Dit antwoord is INCORRECT.
Lachgas is lachgas en geen stikstofmonoxide. Het is niet reactief en niet giftig, maar tast wel de eigenschappen van membranen aan.E. Is niet giftig voor cellen.
Nitrisch Oxide:
A. Heeft een lange levensduur
B. Is membraan doorlaatbaar
C. Is niet reactief
D. Is ook bekend als lachgas.
E. Is niet giftig voor cellen. Dit antwoord is INCORRECT.
NO is zeer giftig voor cellen wanneer het in overmaat wordt geproduceerd. In feite wordt gedacht dat overproductie van NO bepaalde neurodegeneratieve ziekten veroorzaakt. NO wordt ook geproduceerd door macrofagen als het toxische product om binnendringende bacteriën te doden.