Toll-like receptors zijn microbieel-sensor eiwitten die een breed scala van pathogenen herkennen, zij zijn een kritisch eiwit in de eerste bescherming tegen infectie; het genereren van een onmiddellijke maar relatief niet-specifieke respons. Deze mini-review belicht de rol die TLR spelen in de aangeboren immuniteit, met inbegrip van informatie over TLR-signaalroutes en de moleculen die ermee geassocieerd zijn.
Bekijk onze volledige reeks TLR-antilichamen

Toll-like Receptors (TLRs) Mini-review
Download als PDF
TLRs and Innate Immunity
Toll-like receptors (TLRs) zijn essentiële componenten van de aangeboren immuunrespons en zijn daarom een van de sleutelfactoren die betrokken zijn bij de herkenning van en de verdediging tegen binnendringende ziekteverwekkers. Het zijn transmembraanreceptoren die zowel op het celoppervlak als in endosomale vesikels gelokaliseerd kunnen worden. Zij behoren tot de familie van de patroonherkenningsreceptoren (PRR), samen met de mannose- (CD206), NOD- en NALP-receptoren. Aangezien zij betrokken zijn bij zelf- versus niet-zelf identificatie, wordt ook gedacht dat zij betrokken zijn bij auto-immuunziekten.
Toll-like receptoren vormen een familie van eiwitten die essentieel zijn voor het aangeboren immuunsysteem en die geconserveerd zijn van planten tot zoogdieren.
In 1996 en 1997 toonden Lamaitre en collega’s aan dat het gen Toll, een dorsoventrale regulator van de Drosophila ontwikkeling, ook betrokken was bij de aangeboren immuunrespons op schimmels en grampositieve bacteriën (1, 2). Deze rol was consistent met de structurele overeenkomst van het intracellulaire domein van Toll met de interleukine-1B receptor.
Al bijna op hetzelfde moment, kloonde Janeway’s groep een van de zoogdier homologs van Drosophila Toll, later Toll-like receptor 4 (TLR4) genoemd. Zij stelden vast dat een dominant actieve vorm van TLR4 in staat was om NFkB en NFkB-afhankelijke ontstekingsgenen te stimuleren, en dat TLR4 de expressie van CD80 induceert (3).
Expressie van de CD80 co-stimulatoire molecule is noodzakelijk voor de activatie van naïeve T lymfocyten, en de inductie ervan door TLR4 was het eerste bewijs dat het aangeboren immuunsysteem zou kunnen deelnemen aan de polarisatie van naïeve T lymfocyten en adaptieve immuniteit zou kunnen activeren (3).
TLR Genen en Expressie
Tot nu toe zijn 11 TLR genen geïdentificeerd in de mens, en 13 in de muis. Ze kunnen in klassen worden verdeeld op basis van hun functie en sequentie. Hoewel het allemaal membraaneiwitten zijn, zijn sommige TLR’s gelokaliseerd in intracellulaire vesikels.
Hoewel de expressie van TLR’s zeer breed is, met verschillende weefsels die meer dan één type TLR tot expressie brengen, ligt het hoogste expressieniveau en het grootste bereik van TLR’s op die weefsels die betrokken zijn bij aangeboren immuniteit (4).
TLR-geassocieerde Moleculen
Het intracellulaire domein van de TLR’s wordt Toll/IL-1 (TIR) genoemd, wegens overeenkomsten met de IL-1-receptor, terwijl het extracellulaire deel een leucinerijk repeat (LRR)-domein bevat. Van het TIR is bekend dat het betrokken is bij signaaltransductie, terwijl van het LRR wordt verondersteld dat het direct het cognate ligand herkent.
Na binding van het ligand dimeren TLR’s of associëren ze zich met andere receptoren. Het TIR-domein staat in wisselwerking met andere TIR-bevattende intracellulaire eiwitten.
Er zijn vier intracellulaire adaptoreiwitten beschreven die een TIR-domein bevatten:
- MyD88
- TRIF (of TICAM-1)
- TIRAP (of MAL)
- TICAM-2 (of TRAM)
MyD88 en TRIF definiëren twee onafhankelijke signaalpaden, terwijl MAL en TICAM als brugmoleculen fungeren.
TLR-signaalroute
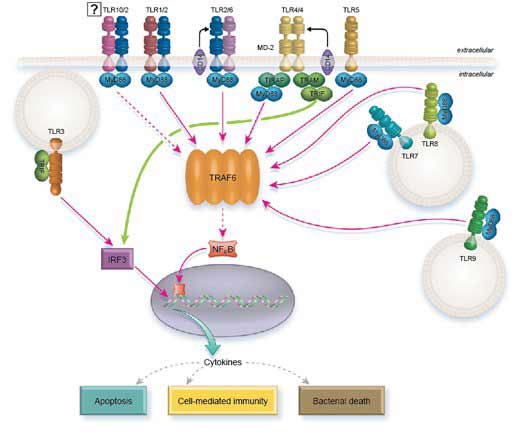
TLR-signaalroute
Alle TLR’s, behalve TLR3, kunnen via MyD88 signaleren. TLR2 en TLR4 hebben echter TIRAP nodig om MyD88 te rekruteren. MyD88 recruteert op zijn beurt IRAK1 en IRAK4 die van MyD88 dissociëren na fosforylering. Vervolgens interageren IRAK1 en IRAK4 met TRAF6, dat op zijn beurt de NFkB-route, AP1 en IRF5 activeert, waardoor uiteindelijk de transcriptie van ontstekingsbevorderende cytokinen wordt gestimuleerd.
Om deze taak te voltooien, fosforyleert TRAF6 IRF5 direct, terwijl het AP1 activeert via MAP-kinases. De vrijlating van NFkB van zijn IKB-remmer vereist de rekrutering van TAK1- en TAB-eiwitten van TRAF6 die op IKB-kinase (IKKa/IKKB/NEMO) inwerken.
TLR7, TLR8, en TLR9 induceren type I IFN-signalering via MyD88. In dit geval wordt MyD88 verondersteld IRF1 direct te activeren en IRF7 indirect te activeren via de interactie van TRAF6 met TRAF3.
In plaats van via MyD88 te werken, signaleert TLR3 via TRIF. De route activeert echter uiteindelijk dezelfde effectoren: NFkB, AP1, IRF3, en IRF7.
TLR4 is uniek omdat het zowel MyD88 als TRIF als intracellulaire adaptor moleculen gebruikt. Het heeft ook het brugmolecuul TRAM nodig om TRIF te rekruteren en TIRAP om MyD88 te rekruteren.
Interessant is dat het blijkt dat TLR4 beide pathways sequentieel gebruikt en niet vanuit hetzelfde compartiment (5). Wanneer TLR4 LPS bindt, wordt TIRAP gerekruteerd en rekruteert op zijn beurt MyD88 signalering vanuit het plasmamembraan om cytokine transcriptie te activeren. LPS bevordert ook de internalisatie van TLR4 door endocytose, een gebeurtenis die MyD88 signalering uitschakelt. Op dat ogenblik blijft de geïnternaliseerde TLR4 signalen geven vanuit het endolysosoom compartiment, maar nu werkt het via TRAM/TRIF en leidt het tot de transcriptie van interferon genen.
Voor meer informatie over patroonherkenningsreceptoren met inbegrip van TLR’s bezoek onze Patroonherkenningsreceptor webpagina en bekijk het gamma beschikbare antilichamen.
- Lemaitre, B. et al. (1996) The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults.
Cell. 86:973-83 - Lemaitre, B. et al. (1997) Drosophila host defense: differential induction of antimicrobial peptide genes after infection by various classes of microorganisms.
Proc Natl Acad Sci U S A. 94:14614-9 - Medzhitov, R. et al. (1997) A human homologue of the Drosophila Toll protein signals activation of adaptive immunity.
Nature. 388:394-7 - Zarember, K.A. and Godowski, P.J. (2002) Tissue expression of human Toll-like receptors and differential regulation of Toll-like receptor mRNAs in leukocytes in response to microbes, their products, and cytokines.
J. Immunol. 168:554-561 - Barton, G.M. and Kagan, J.C. (2009) A cell biological view of Toll-like receptor function: regulation through compartmentalization.
Nat Rev Immunol. 9:535-42
Verder lezen
- Montero Vega, M.T. and De Andrés Martín, A. (2009) The significance of toll-like receptors in human diseases.
Allergol Immunopathol(Madr). 37:252-63 - Montero Vega, M.T. and De Andrés Martín, A. (2008) Toll-like receptors: a family of innate sensors of danger that alert and drive immunity.
Allergol Immunopathol(Madr). 36:347-57 - Uematsu, S. and Akira, S. (2008) Toll-Like receptors (TLRs) and their ligands.
Handb Exp Pharmacol. :1-20 - Bauer, S. et al. (2009) Pattern recognition by Toll-like receptors.
Adv Exp Med Biol. 653:15-34 - Schmidt M. et al. (2010) Cruciale rol voor humane Toll-like receptor 4 bij de ontwikkeling van contactallergie voor nikkel.
Nat Immunol. 2010 Aug 15.