GPIアンカー型タンパク質は変わり者です。 細胞生物学の入門書では、膜タンパク質は5種類あり、次のような名前がついていると教わりました。 タイプI、タイプII、タイプIII、タイプIV、そしてGPIアンカー型です。 なぜ、このような奇妙なタンパク質が、糖鎖と脂肪鎖に融合しているのでしょうか? 彼らは何をしているのでしょうか?
ソニアと私、そしてチームメイトのアンドリューは、このテーマについていくつかの文献を読み、学んだことを共有するためにこのブログ記事を書いています。 これらは主に、驚くべきことが知られている GPI アンカー自体の構造と生合成をカバーしています。
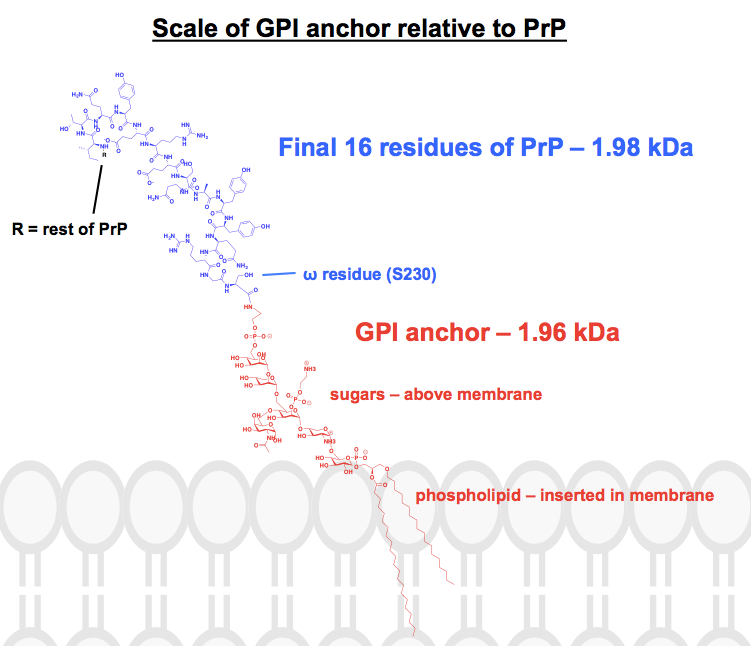
このアンカー(正式名称はグリコシルホスファチジルイノシトール)は一枚岩ではなく、詳細はさまざまである分子の一般的な記述です。 一般的には、タンパク質のω(翻訳後の最後の)残基から始まって、エタノールアミン、リン酸、いくつかの糖、そしてリン脂質がある。 核となる糖骨格は保存されているが、そこから分岐する側鎖は様々であり、リン脂質のヘッドグループと脂肪酸も同様に変化しうるのである。 PrP の GPI アンカーは、1994 年に特徴づけられましたが、それでも一枚岩ではなく、糖側鎖の組成が異なる少なくとも 6 種類の構造が確認されました。 私は、これらのアンカーが化学的にどのようなもので、付着したタンパク質との関連において実際にどのようなものであるかを知りたかったので、ChemDraw で実際に完全な構造を描いてみることにしました。 図1のうち、最も完全な骨格構造に近いものを参考にしながら、図6の上部にあるPrPのGPIアンカーの詳細を書き加えていきました。 分子量は1,958 Daであったので、文脈からHuPrP23-230の最後の16残基を描き加えてみたところ、1,979 Daとなり、同等の重さとなった。 これは、PrPの翻訳後修飾配列の約8%にあたる。 すべての結合が正しいかどうかはわかりませんが、以下は私が思いついたものです:
多くの場合、遺伝子は複数のアイソフォームを持ち、あるスプライシング生成物は GPI アンカータンパク質を生じ、他のものは分泌型または膜貫通型を生じさせる。 例えば、NCAM1はGPIアンカー型と膜貫通型の2つのアイソフォームを持ち、ACHE(アセチルコリンエステラーゼをコード)はGPIアンカー型が赤血球上にのみ存在する(NCBI Genes)。 ここで最も興味深いのは、マウスの遺伝子Ly6aの話である。この遺伝子は、遺伝子多型のおかげで、ある系統のマウスではGPIアンカー型になり、他の系統ではならないのである。 そのGPIアンカー型でのみ、ウイルスベクターAAV PHP.eB の受容体として働きます(このベクターは、遺伝子治療のために脳神経細胞に驚くほど効率的に取り込まれますが、悲しいことに、それはマウスだけの遺伝子で、私たち人間はLy6aを持ってさえいません)。
GPIアンカーがどのように合成され、タンパク質に結合するかについては多くのことが知られており、>20個のタンパク質がこの経路に関与し、そのほとんどは接頭辞「PIG」で始まり、PIGA、PIGKなどの遺伝子によりコードされています(図2参照)。 生合成のほとんどは、アンカーが小胞体内の膜に挿入された状態で行われるが、どのタンパク質にも結合していない。 実際、最初の数段階は膜の細胞質側のリーフレットで行われ、後になってアンカーが内腔側(ER内)に反転するのである。 最終的には、少なくとも5つのタンパク質からなる複合体であるGPIトランスアミダーゼが、タンパク質のC末端からGPIシグナルを切断し、タンパク質のいわゆるω残基(翻訳後修飾配列の最後の残基)にGPIアンカーを付着させることで完結する。
真菌におけるGPI生合成の低分子阻害剤は数多く存在し、そのいくつかは抗真菌薬として開発されようとしているが、私が知る限り、哺乳類細胞におけるGPI生合成阻害剤は、マンノサミン(マンノース類似体で、GPIへの取り込みに化学的に不適合)しか知られていない … 続きを読む
GPIトランスアミダーゼがどのようなアミノ酸配列モチーフを認識するのか、配列ロゴを探してみましたが、見つかりませんでした。 どうやら配列モチーフはかなり緩いようで、どうやらGPIシグナルは相同ですらない、つまり共通の祖先配列から進化したのではなく、収束さえして進化しているようです。 私が見つけた最も良い説明は、(N-C-末端をタンパク質の末端まで読むと)1)約11残基の非構造化リンカー、2)S、N、D、G、A、Cのいずれかのω残基を含む小さな側鎖を持つ数残基、3)5-10個の極性アミノ酸のスペーサー、最後に4)15-20個の疎水性アミノ酸が必要だというものであった。 PrPはこのモチーフを緩やかに踏襲している。 公表されている構造によると、αヘリックス3はQ223残基で終わっており、「非構造化リンカー」はちょうどAYYQR(規定の11残基よりやや短い)になっている。 小さな側鎖」領域はGS|SM(パイプはトランスアミダーゼ切断部位を示す)、極性領域はVLFSSPP、疎水性C末端はVILLISFLIFLIVGであろう。
GPI生合成・付着経路のタンパク質のいくつかは非常に重要であり、PIGO、PIGV、PIGW、PGAP2、PGAP3などの遺伝子における二重機能喪失型または明らかに低型のミスセンス変異によるGPIアンカー欠損の多くの重症の疾患や症候が報告されている(By blible loss-of-function or apparently hypomorphic missense mutation in the genes like PIGO, PIGV, PIGW, PGAP2, and PGAP3 )。
ソニアは、2つのGPIアンカータンパク質の生合成に必要な遺伝子を特定するために、ハプロイドヒト細胞で突然変異誘発スクリーニングを行った数年前の優れた論文を見つけました。 PrPとCD59です。 彼らは、細胞表面の PrP と CD59 に基づいて細胞の FACS ソーティングを繰り返し、これらのタンパク質の表面レベルが劇的に低下した細胞を特定し、次に配列決定を行い、これらの細胞で親集団に対してどの遺伝子ノックアウトが濃縮されているかを確認しました。 予想通り、PIG遺伝子のほとんどが両タンパク質でヒットしましたが(図4)、すべてのヒットが重なっているわけではありません。特に、RNAレベルでは、PrPとCD59は組織間で最も類似した発現プロファイルを持つタンパク質なので、これは少し驚きです(この記事の一番下にある熱マップを参照)。 GPIアンカーの側鎖修飾に関与する多くの酵素がCD59でのみ見つかったことから、PrPではなくCD59が成熟して細胞表面に到達するためには、これらの複雑な側鎖が必要であることが示唆される。 一方、Sec62とSec63はPrPに対してのみ検出された。これらはERへの翻訳共役型トランスロケーションに何らかの形で関与するタンパク質だが、PrPには必要だがCD59やCD55、CD109といった他のコントロールタンパク質には必要ないことが明らかにされた。 このことは、「PrPの発現には何か特別なものがあるのか」という私の疑問に対する答えとして、魅力的な新章を開くものである。 もちろん、これらのタンパク質が他の3つの制御タンパク質にとって重要でなかったからと言って、重要でないということにはなりません。ある研究では、Sec62が多くの小タンパク質の分泌に必要であることがわかりましたし、SEC62遺伝子は、ヒト集団において機能喪失型変異が完全に欠落しており、ハプロ不全を示唆するに十分な数となっています。 SEC63はあまり制約を受けていないように見えますが、それは単に劣性的に作用しているということかもしれません。
上記のいずれも、なぜGPIアンカータンパク質が存在するのかという疑問に対する答えにはなっていません。 ところで、私が昔受けた細胞生物学の授業では、詳細が省略されていました。実は膜タンパク質には、テールアンカーデッド(TA)タンパク質と呼ばれる、疎水性のC末端を持ち、膜に刺さるが反対側には突き出さない6番目のクラスがあるのです。 なぜ、GPIアンカータンパク質はすべてTAタンパク質になれなかったのでしょうか? なぜ細胞は、代わりに糖-脂肪アンカーを合成するような複雑な経路を進化させたのでしょうか。また、なぜこのように早い段階で進化させたのでしょうか。 GPIアンカータンパク質自体は、その本来の機能が知られている範囲では、酵素(AChEなど)、細胞接着分子(NCAM1など)、免疫系で補体を制御するタンパク質(CD59)など、非常に幅広い機能を有しているのだが . 末梢神経のミエリン維持に関与するGPIアンカータンパク質は、少なくとも1つは存在するらしい。 しかし、GPIアンカータンパク質は、他のタンパク質にはできない、いったいどんなことができるのだろうか? ある総説では、これまで提案されたいくつかのアイデアを挙げている。 一つは、GPIアンカータンパク質は一過性の二量体形成に長けているというものである。 いくつかの研究では、プリオンの生物学において二量体化が何らかの役割を担っているという考えが示されているが、そこで用いられたモデル系と生体内の状況との関連はまだ明らかではない。 もう一つの考え方は、GPIアンカータンパク質は、例えばアンジオテンシン変換酵素(ACE)によって細胞表面から排出されることがあるので、その局在は何らかの動的な方法で制御されうるというものである。 この点でも、PrPはADAM10という酵素によって排出されることが分かっているが、PrP本来の機能にどのような役割を果たしているかはまだ明らかではない。 第三のアイデアは、GPIアンカータンパク質が「脂質ラフト」に選択的に集まるというもので、おそらく私が最もよく耳にするものです。 なぜなら、これらのタンパク質の局所的な有効濃度が高まることで、より多くの相互作用が可能になるなど、あらゆる種類のノックオン効果が想像されるからです。 脂質ラフトは、機能的には界面活性剤の不溶性によって定義され、多くの人がスフィンゴミエリンとコレステロールに富むと表現しますが、何が脂質ラフトで、何が脂質ラフトではないのか、普遍的に受け入れられる定義はありませんし、経験則から、多くの人が考えるよりずっと小さく、もっと一時的かもしれない、と指摘している人もいます。
そこで、これらのタンパク質のリストを入手し、それらがどのようなものであるかをよりよく理解するために、いくつかの分析を行いました。 これらは 140 の遺伝子シンボルにマップされますが、このスクリプトを実行して現在 HGNC で承認されているタンパク質をコードする遺伝子シンボルに更新すると、135 に減少します。 859>
Uniprot は、そのアノテーションがどのように生成されたかについての情報を提供していませんが、かなりの程度、手作業によるキュレーションが行われているはずです。 比較のために、Andrewは、GPIアンカーを切断する2つの酵素であるPI-PLDまたはPI-PLCを用いて、細胞からGPIアンカータンパク質を経験的に分離した一連の素晴らしい論文も掘り起こしました。 これらの論文のリストを組み合わせて、現在の遺伝子記号にマッピングすると、107の遺伝子が得られた。 この中からランダムに数個を抜き出し、スポットチェックを行った。 その中には、グリピカン-1 (GPC1) や神経細胞接着分子 (NCAM1) などのよく知られたGPIアンカータンパク質が含まれており、これらはいずれもPrPと相互作用することが報告されている。 しかし、VDAC3のように、文献上GPIアンカーが知られていないと思われる遺伝子もいくつか存在した。これらの中には、単に非常に豊富なタンパク質であるか、他の理由による偽陽性である可能性もある。 一方、偽陰性の明らかな原因として、調査した細胞株では単に発現していない遺伝子や、質量分析で拾い上げるほど豊富でない遺伝子、PrPパラログのSPRNとPRNDがリストに含まれていないことなどがある。 全体として、51の遺伝子が両方のリストにあり、非常に有意な濃縮(OR = 217, P < 1 × 10-84)であり、Uniprotのアノテーションが経験的データと一致することを再確認することができた。
このリストで武装して、GPIアンカータンパク質がどのように積み重なるかを見てみたいと思いました。 PrPはシングルエクソンで、短く(成熟型で208アミノ酸)、非必須で、広く発現しているタンパク質である。 GPI アンカータンパク質は、他のタンパク質と同様に、あらゆる次元で多様であることがわかりました。 以下は、全遺伝子とGPIアンカータンパク質をコードする遺伝子のコーディング配列の長さを塩基対で重ね合わせたヒストグラムです。 GPIアンカー型はぎりぎり左にシフトした分布になっている。 GPIアンカータンパク質遺伝子の平均的なコード配列長は1,301 bpであり、平均的な遺伝子は1,729であるが、この平均値の差は各グループ内の変動に比べれば小さいものである。 CD52は配列がわずか186塩基対で、成熟型ではわずか12アミノ酸であり、GPIアンカータンパク質の中では最小である。 GPIアンカータンパク質は全遺伝子と比較して、平均してエクソンがやや少なく(平均7.8対10.1)、上述のわずかな長さの分布の違いと一致するが、ほとんどはマルチエクソンである。 ここでもPrPは小さい方である。1つのコーディングエキソンしか持たないGPIアンカータンパク質は6つしかなく、そのうちの3つがPrPとその2つのパラログであるShoとDplである。 (他の 3 つの遺伝子は GAS1、SPACA4、そして素晴らしい名前の OMG です)
次に、機能喪失の制約について調べました。 制約とは、例えばナンセンス、フレームシフト、スプライスサイトの変異が、突然変異率に基づく予想と比較して、一般集団でどれだけ枯渇しているかに基づいて、遺伝子がどれだけ強い自然選択を受けているかを示す尺度です。 この指標は、統計的な理由(短い遺伝子では予想される突然変異の数が少ないので、枯渇の定量化が難しい)と生物学的な理由(シングルエクソン遺伝子はナンセンスを介した崩壊を受けないので、タンパク質切断変異が本当に「機能喪失」であるかどうかを知ることが難しい)の両方により、短い遺伝子ではあまり解釈の難しい指標である。 しかし、ほとんどのGPIアンカータンパク質はPrPほど短くないので、見てみる価値があると思った。 その結果、平均して、GPIアンカータンパク質は、平均的な遺伝子よりもわずかに制約が少ない、つまり、機能喪失変異の期待される量をより多く持っていることがわかりました。 平均的な遺伝子は機能喪失変異の47%を持っており、GPIアンカータンパク質は56%を持っているのである。 しかし、ここでのすべてと同様に、どちらの陣営にも広い分布があります。 GPIアンカータンパク質の場合、絶対的な制約を受けるACHE(17のLoFが予想され、観測されなかった)と、もう一方の端には、機能喪失に対して全く選択されないように見えるいくつかの遺伝子(CNTN6、CD109、TREH、MSLNがいくつかの例です)があるのです。 PRNPは、タンパク質切断変異体が機能獲得を引き起こす≧145の残基を除外すると、後者の陣営に入る。
最後に、GPIアンカータンパク質はどこで発現しているのだろうかと考えた。 PRNPは脳で最も多く発現しているが、どこでも発現している。 これは典型的な例でしょうか。 GTEx v7 “gene median tpm” summary file (Jan 15, 2016)をフルダウンロードしました。行ごとに遺伝子、列ごとに組織、セルはRPKM – RNA-seq reads per kilobase of exon per million mapped readsとなっています。 このデータセットで作業するには、いくつかの工夫が必要でした。 バイオインフォマティシャンの中には、<1 RPKMを「発現していない」とみなす人もいると聞きますが、発現行列はまばらで、ほとんどの遺伝子はほとんどの組織で高発現しないため、生のRPKMをプロットしただけでは、1 RPKM以下のノイズが支配的になる可能性があるのです。 一方、遺伝子発現は対数スケールで考える必要があります。組織内の遺伝子は<1RPKMから>10,000RPKMまで変化するため、すべてを線形スケールで考えると、少数の本当に高発現の遺伝子と組織の組み合わせも支配的になり、行列は実際よりもさらにまばらなものになってしまうのです。 そこで、行列のlog10をとって、分布を , で切り捨てて、紫色のスケールで、1 – 10 – 100 – 1,000 – 10,000 RPKMを実行しました。 それから、UniprotのGPIアンカータンパク質にサブセットしました。 これを可視化するために、生まれて初めてヒートマップを作りました。 論文などでよく目にするヒートマップですが、普段はあまりピンとこないのですが、今回は発現のパターンを知ることが目的でしたので、少し遊んでみたところ、これが一番ピンとくるものでした。 ヒートマップの原理は、行と列が、似たようなものが一緒になるようにクラスタリングされることです。 例えば、脳組織の列はx軸のパッチに連続して並び、脳で高発現している遺伝子はy軸のパッチに連続して並び、それらの交点が濃い紫色の長方形を形成することで、「ほとんど脳に発現している遺伝子群が存在する」と解釈できます。
興味のある読者は、ヒートマップのフルスケールのベクターアート PDF を見ることができますが、よりすぐにアクセスできるように、関心のあるクラスターを呼び出した手書き注釈版を以下に示します。 PRNPは、CD59、LY6E、GPC1、BST2とともに、このヒートマップの最上部に位置する、より高度で広範に発現する一握りのものの1つです。 ほとんどのGPIアンカータンパク質は、より低い、あるいはより組織に限定された発現をしており、あるものはほとんど脳にのみ発現し、あるものはほとんど脳には発現しない。また、他の小さなクラスタは、PrPのパラログPRNDのように精巣などの特定の組織に主に属し、ノックアウトにより男性不妊症を引き起こす。
結論
GPIアンカータンパク質は、その機能が知られている限り、ほぼすべてのサイズで、ほぼすべての組織で発現し、ほぼすべての機能を有するようです。 多くのGPIアンカータンパク質は非常に明確な本来の機能を持っているが、これらの機能は多様であり、特にこれらのタンパク質の多くは非GPIアンカーのアイソフォームも存在するため、なぜGPIアンカーを必要とするのかは明らかでない。 一方、PrPを含む他のGPIアンカー型タンパク質については、そもそもネイティブな機能について十分に分かっていないため、なぜネイティブな機能がGPIアンカーを必要とするのかを推測することすら困難である。 私が行った解析や読んだレビューのどれもが、なぜこのアンカー機構が存在するのか、何がこれらのタンパク質にそれを必要とさせているのか、統一的な原理を導き出すことはできませんでした。 GPIアンカーを持つタンパク質がなぜユニークなのかについては、脂質ラフト、ホモダイマー、シェディングなど、いくつかの仮説があります。 これらの仮説はすべて、ある程度の説得力を持つかもしれない。
この投稿の解析のためのRコードと生データファイルはこちらです
。