I recettori Toll-like sono proteine di rilevamento microbico che riconoscono una vasta gamma di patogeni, sono una proteina critica nella protezione iniziale contro l’infezione, generando una risposta immediata ma relativamente aspecifica. Questa mini-review evidenzia il ruolo che i TLR svolgono nell’immunità innata, comprese le informazioni sulle vie di segnalazione dei TLR e le molecole ad essi associate.
Visualizza la nostra gamma completa di anticorpi TLR

Recettori simil-Toll (TLR) Mini-review
Scarica come PDF
TLR e immunità innata
I recettori simil-Toll (TLR) sono componenti essenziali della risposta immunitaria innata e sono quindi uno dei fattori chiave coinvolti nel riconoscimento e nella difesa contro agenti patogeni invasori. Sono recettori transmembrana che possono essere localizzati sia sulla superficie cellulare che su vescicole endosomiali. Appartengono alla famiglia dei pattern recognition receptor (PRR) insieme ai recettori del mannosio (CD206), NOD e NALP. Dal momento che sono coinvolti nell’identificazione del sé contro il non sé, si pensa che siano anche implicati nelle malattie autoimmuni.
I recettori simil-Toll costituiscono una famiglia di proteine essenziali per il sistema immunitario innato che è conservata dalle piante ai mammiferi.
Nel 1996 e 1997, Lamaitre e colleghi hanno dimostrato che il gene Toll, un regolatore dorsoventrale dello sviluppo della Drosophila, era anche coinvolto nella risposta immunitaria innata ai funghi e ai batteri gram-positivi (1, 2). Questo ruolo era coerente con la somiglianza strutturale del dominio intracellulare di Toll al recettore dell’interleuchina-1B.
Quasi allo stesso tempo, il gruppo di Janeway ha clonato uno degli omologhi mammiferi di Drosophila Toll, poi chiamato Toll-like receptor 4 (TLR4). Hanno osservato che una forma attiva dominante di TLR4 era in grado di stimolare NFkB e i geni infiammatori dipendenti da NFkB, e che TLR4 induce l’espressione di CD80 (3).
L’espressione della molecola co-stimolatoria CD80 è necessaria per l’attivazione dei linfociti T ingenui, e la sua induzione da parte di TLR4 è stata la prima prova che il sistema immunitario innato potrebbe partecipare alla polarizzazione dei linfociti T ingenui e attivare l’immunità adattativa (3).
Geni TLR ed espressione
Sino ad ora 11 geni TLR sono stati identificati nell’uomo e 13 nel topo. Possono essere divisi in classi in base alla loro funzione e sequenza. Anche se tutti sono proteine di membrana, alcuni TLR sono localizzati in vescicole intracellulari.
Mentre l’espressione dei TLR è molto ampia, con diversi tessuti che esprimono più di un tipo di TLR, il livello di espressione più alto e la gamma più ampia di TLR si trovano in quei tessuti coinvolti nell’immunità innata (4).
Molecole associate ai TLR
Il dominio intracellulare dei TLR è denominato Toll/IL-1 (TIR), a causa delle somiglianze con il recettore IL-1, mentre la parte extracellulare contiene un dominio LRR (leucin-rich repeat). Il TIR è noto per essere coinvolto nella trasduzione del segnale, mentre il LRR è ritenuto riconoscere direttamente il ligando cognato.
In seguito al legame con il ligando, i TLR si dimerizzano o si associano con altri recettori. Il dominio TIR interagisce con altre proteine intracellulari contenenti TIR.
Sono state descritte quattro proteine adattatrici intracellulari che contengono un dominio TIR:
- MyD88
- TRIF (o TICAM-1)
- TIRAP (o MAL)
- TICAM-2 (o TRAM)
MyD88 e TRIF definiscono due vie di segnalazione indipendenti, mentre MAL e TICAM agiscono come molecole ponte.
Percorso di segnalazione TLR
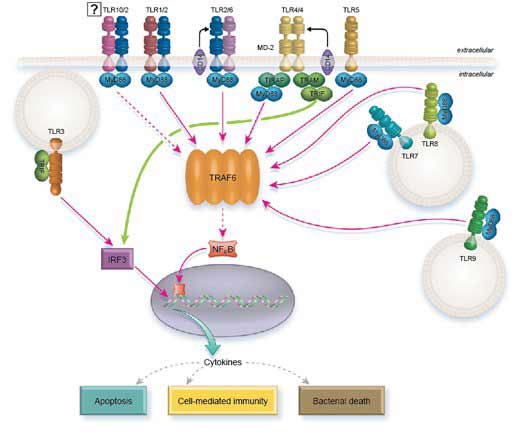
Segnalazione TLR
Tutti i TLR, tranne TLR3, possono segnalare attraverso MyD88. Tuttavia, TLR2 e TLR4 richiedono TIRAP per reclutare MyD88. A sua volta, MyD88 recluta IRAK1 e IRAK4 che si dissociano da MyD88 dopo la fosforilazione. Successivamente, IRAK1 e IRAK4 interagiscono con TRAF6 che a sua volta attiva la via NFkB, AP1 e IRF5, stimolando infine la trascrizione delle citochine infiammatorie.
Per completare questo compito, TRAF6 fosforila direttamente IRF5, mentre attiva AP1 attraverso le MAP chinasi. Il rilascio di NFkB dal suo inibitore IKB richiede il reclutamento delle proteine TAK1 e TAB di TRAF6 che agiscono sulla chinasi IKB (IKKa/IKKB/NEMO).
TLR7, TLR8, e TLR9 inducono la segnalazione di IFN di tipo I attraverso MyD88. In questo caso si ritiene che MyD88 attivi direttamente IRF1 e indirettamente IRF7 attraverso l’interazione di TRAF6 con TRAF3.
Invece di agire attraverso MyD88, TLR3 segnala attraverso TRIF. Tuttavia, il percorso finisce per attivare gli stessi effettori: NFkB, AP1, IRF3 e IRF7.
TLR4 è unico in quanto utilizza sia MyD88 che TRIF come molecole adattatrici intracellulari. Richiede anche la molecola ponte TRAM per reclutare TRIF e TIRAP per reclutare MyD88.
Interessante, sembra che TLR4 usi entrambi i percorsi in modo sequenziale e non dallo stesso compartimento (5). Quando TLR4 si lega a LPS, TIRAP viene reclutato e a sua volta recluta la segnalazione MyD88 dalla membrana plasmatica per attivare la trascrizione delle citochine. LPS promuove anche l’internalizzazione di TLR4 attraverso l’endocitosi, un evento che spegne la segnalazione di MyD88. A questo punto il TLR4 internalizzato continua a segnalare dal compartimento dell’endolisosoma, ma ora agisce attraverso TRAM/TRIF e porta alla trascrizione dei geni dell’interferone.
Per ulteriori informazioni sui recettori di riconoscimento del modello, compresi i TLR, visita la nostra pagina web sui recettori di riconoscimento del modello e rivedi la gamma di anticorpi disponibili.
- Lemaitre, B. et al. (1996) La cassetta gene regolatore dorsoventrale spätzle/Toll/cactus controlla la potente risposta antifungina in Drosophila adulti.
Cell. 86:973-83 - Lemaitre, B. et al. (1997) Drosophila host defense: induzione differenziale dei geni peptidi antimicrobici dopo l’infezione da varie classi di microrganismi.
Proc Natl Acad Sci U S A. 94:14614-9 - Medzhitov, R. et al. (1997) Un omologo umano della proteina Toll di Drosophila segnala l’attivazione dell’immunità adattativa. 388:394-7
- Zarember, K.A. and Godowski, P.J. (2002) Espressione tissutale dei recettori Toll-like umani e regolazione differenziale degli mRNA dei recettori Toll-like nei leucociti in risposta a microbi, loro prodotti e citochine.
J. Immunol. 168:554-561 - Barton, G.M. and Kagan, J.C. (2009) A cell biological view of Toll-like receptor function: regulation through compartmentalization.
Nat Rev Immunol. 9:535-42
Altra lettura
- Montero Vega, M.T. and De Andrés Martín, A. (2009) The significance of toll-like receptors in human diseases.
Allergol Immunopathol(Madr). 37:252-63 - Montero Vega, M.T. and De Andrés Martín, A. (2008) Toll-like receptors: a family of innate sensors of danger that alert and drive immunity.
Allergol Immunopathol(Madr). 36:347-57 - Uematsu, S. and Akira, S. (2008) Toll-Like receptors (TLRs) and their ligands.
Handb Exp Pharmacol. :1-20 - Bauer, S. et al. (2009) Pattern recognition by Toll-like receptors.
Adv Exp Med Biol. 653:15-34 - Schmidt M. et al (2010) Crucial role for human Toll-like receptor 4 in the development of contact allergy to nickel.
Nat Immunol. 2010 Aug 15.