A GPI-horgonyzott fehérjék a különcök közé tartoznak. A bevezető sejtbiológiában azt tanították nekünk, hogy a membránfehérjéknek öt típusa van, amelyeket a következőképpen neveztek el: I. típusú, II. típusú, III. típusú, IV. típusú és GPI-horgonyzott. Miért van a fehérjéknek ez a furcsa, cukor-zsírlánccal összefűzött osztálya? Mi a feladatuk? Kaphatunk-e valamilyen betekintést az engem érdeklő fehérjébe – a PrP-be -, ha többet tudunk meg erről a fehérjeosztályról, amelynek tagja?
Sonia és én, valamint Andrew csapattársunk és némi olvasmányt végeztünk ebben a témában, és azért írom ezt a blogbejegyzést, hogy megosszunk néhányat abból, amit megtudtunk.
olvasás
Azzal kezdtük, hogy elolvastunk néhány áttekintést . Ezek leginkább magának a GPI horgony szerkezetével és biogenezisével foglalkoztak, amelyről elképesztő dolgokat tudunk.
Ez a horgony, amelynek teljes neve glikozil-foszfatidil-inozitol, nem egy monolit: ez egy olyan molekula általános leírása, amelynek részletei változhatnak. Általánosságban elmondható, hogy a fehérje ω (utolsó poszttranszlációsan jelenlévő) maradékától kezdve van egy etanolamin, majd egy foszfát, majd néhány cukor, végül egy foszfolipid. A magcukor gerinc konzerválódik, de a belőle elágazó oldalláncok változhatnak, és a foszfolipid fejcsoport és a zsírsavak is változhatnak. A PrP GPI horgonyát a , de még akkor sem egy monolit – legalább hat különböző struktúrát azonosítottak, amelyek különböznek a cukor oldalláncok összetételében.
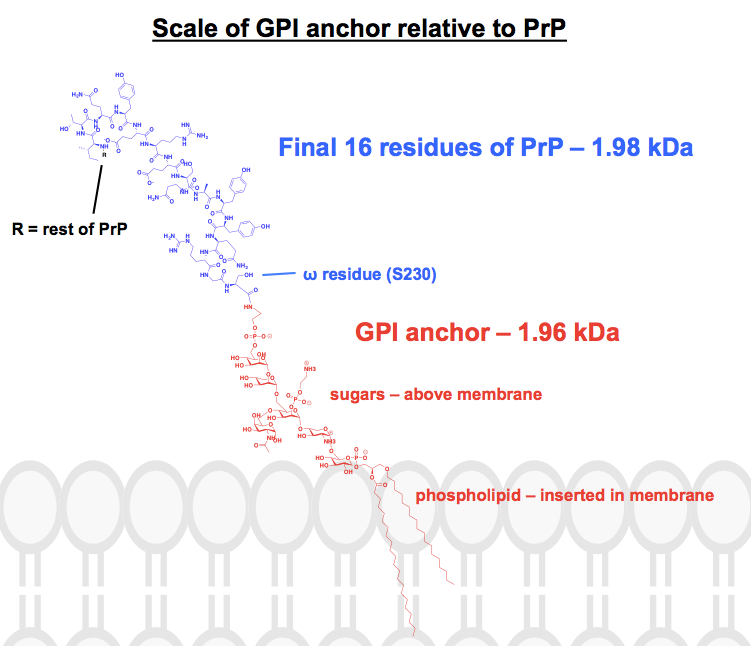
Minden kémiai szerkezet, amit a GPI horgonyokról találtam, legalább néhány részét rövidítve vagy összefoglalva tartalmazza, és a fehérje általában csak képként van feltüntetve. Szerettem volna képet kapni arról, hogy ezek a horgonyok valójában hogyan néznek ki kémiailag, a hozzájuk kapcsolódó fehérjékkel összefüggésben, ezért nekiláttam, hogy ténylegesen megrajzoljak egy teljes szerkezetet ChemDraw-ban. Az 1. ábráról kiindulva – ami a legközelebb állt egy teljes vázszerkezethez, amit találtam – hozzáadtam a PrP egyik GPI-horgonyának részleteit a 6. ábra felső paneléről. A molekulatömeg 1 958 Da lett, így a kontextus kedvéért berajzoltam a HuPrP23-230 utolsó 16 maradékát, amelyek súlya hasonló, 1 979 Da. Ez a PrP poszt-transzlációsan módosított szekvenciájának körülbelül 8%-a. Nem vagyok biztos benne, hogy minden kötést helyesen írtam le, de a következőket találtam:
Sok esetben egy génnek több izoformája van, az egyik splicingtermék GPI-horgonyzott fehérjét eredményez, míg a többi szekretált vagy transzmembrán formát. Ilyen például az NCAM1, amelynek három fő izoformája van, amelyek közül az egyik GPI-horgonyzott, a másik kettő pedig transzmembrán , és az ACHE (acetilkolinészterázt kódoló), amelynek GPI-horgonyzott formája nyilvánvalóan csak a vörösvértestekben található meg (NCBI Genes). A legizgalmasabb történet itt az egér Ly6a génjének története, amely egy genetikai polimorfizmusnak köszönhetően egyes egértörzsekben GPI-horgonyzott, míg másokban nem. Csak GPI-horgonyzott formájában működik receptorként az AAV PHP.eB vírusvektor számára (ez a vektor elképesztően hatékony felvételt tesz lehetővé az agyi idegsejtekbe génterápiás célokra, de sajnos ez csak egérgén – nekünk, embereknek nincs is Ly6a génünk).
Sokat tudunk arról, hogy a GPI-horgonyok hogyan szintetizálódnak és kapcsolódnak a fehérjékhez , az útvonalban >20 fehérje vesz részt, amelyek többsége a “PIG” előtaggal kezdődik, és olyan gének kódolják, mint a PIGA, PIGK és így tovább – lásd a 2. ábrát a diagramért. A bioszintézis nagy része úgy zajlik, hogy a horgony az ER-ben a membránba illesztve van, de nem kapcsolódik semmilyen fehérjéhez. Valójában az első néhány lépés a membrán citoszolikus szárnyán zajlik, és csak később fordul át a horgony a lumenes oldalra (az ER belsejébe). Az utolsó lépés az, amikor a GPI-transzamidáz, egy legalább öt fehérjéből álló komplex lehasítja a GPI-szignált a fehérje C-terminusáról, és a GPI-horgonyt a fehérje úgynevezett ω-maradékához (a poszttranszlációsan módosított szekvencia utolsó maradékához) csatolja. Ezután a GPI-horgony további érése következik be, ahogy a fehérje kivándorol az ER-ből a sejtfelszín felé.
A GPI-bioszintézisnek számos kismolekulás gátlója létezik a gombákban, amelyek közül néhányat már megpróbáltak gombaellenes gyógyszerként kifejleszteni, de amennyire én tudom, az emlős sejtekben a GPI-bioszintézis egyetlen ismert gátlója a mannozamin, egy mannózanalóg, amely kémiailag nem kompatibilis a GPI-be való beépüléssel.
Nézegettem és kerestem egy szekvencia logót, hogy milyen aminosav szekvencia motívumot ismer fel a GPI transzamidáz, de nem találtam. Úgy tűnik, hogy a szekvencia motívum elég laza , és úgy tűnik, hogy a GPI jelek nem is homológok , ami azt jelenti, hogy nem egy közös ősi szekvenciából fejlődtek ki, hanem inkább konvergens módon fejlődtek, amennyiben egyáltalán van konvergencia. A legjobb leírás, amit találtam, az, hogy (a fehérje végéig N-től C végig olvasva) szükség van 1) kb. 11 maradék strukturálatlan linkerre, 2) néhány maradékra kis oldalláncokkal, beleértve egy ω maradékot, ami lehet S, N, D, G, A vagy C, 3) egy 5-10 poláris aminosavból álló spacerre, és végül 4) 15-20 hidrofób aminosavra. A PrP lazán követi ezt a motívumot. A közzétett szerkezetek szerint az alfa-hélix 3 a Q223 maradéknál ér véget, így a “strukturálatlan linker” csak AYYQR marad (valamivel rövidebb, mint az előírt 11 maradék). A “kis oldalláncú” régió GS|SM (a cső a transzamidáz vágási helyet jelöli), a poláris régió VLFSSPP, a hidrofób C terminus pedig VILLISFLIFLIVG.
A GPI bioszintézis és kötődési útvonal néhány fehérjéje nagyon fontos, és számos súlyos betegséget és GPI-horgonyhiányos szindrómát írtak le, amelyek a PIGO, PIGV, PIGW, PGAP2 és PGAP3 gének biallelikus funkcióvesztéséből vagy nyilvánvalóan hipomorf miszenzmutációjából erednek.
Sonia talált egy néhány évvel ezelőtti kiváló tanulmányt, amelyben haploid emberi sejtekben mutagenezis-szűrést végeztek két GPI-horgonyzott fehérje biogeneziséhez szükséges gének azonosítására: PrP és CD59 . A sejtek ismételt FACS-szortírozását használták a sejtfelszíni PrP és CD59 alapján, hogy azonosítsák azokat a sejteket, amelyekben drámaian csökkent e fehérjék felszíni szintje, majd szekvenálást végeztek, hogy megnézzék, mely gének knockoutjai gazdagodtak ezekben a sejtekben a szülőpopulációhoz képest. Amint az várható volt, a legtöbb PIG gén mindkét fehérje esetében előkerült (4. ábra), de nem minden találat fedte egymást, ami kissé meglepő, különösen azért, mert legalábbis RNS-szinten a PrP és a CD59 a két olyan fehérje, amelyeknek az expressziós profilja a leghasonlóbb a szövetekben (lásd a hőtérképet a bejegyzés alján). Egy csomó, a GPI-horgony oldallánc-módosításában részt vevő enzim csak a CD59 esetében bukkant fel, ami arra utal, hogy a CD59-nek, de a PrP-nek nem, szüksége van ezekre a komplex oldalláncokra ahhoz, hogy megérjen és elérje a sejtfelszínt. Eközben a Sec62 és Sec63 csak a PrP esetében került elő – ezek a fehérjék valamilyen módon részt vesznek az ER-be történő ko-transzlációs transzlokációban, de úgy tűnik, hogy a PrP-nek szüksége van rájuk, de nem a CD59-nek, sem a CD55-nek vagy a CD109-nek, két másik vizsgált kontrollfehérjének. Ez egy lenyűgöző új fejezet a “van-e valami különleges a PrP expressziójában?” kérdésemre adott válaszban, ahol a PrP biogenezisében kerestem valami egyedit, ami potenciálisan megcélozható lenne egy kis molekulával. Természetesen, csak azért, mert ezek a fehérjék nem voltak fontosak három másik kontrollfehérje számára, nem jelenti azt, hogy nem fontosak – egy tanulmány szerint a Sec62 szükséges számos kis fehérje szekréciójához , és a SEC62 gén teljesen kimerül a loss-of-function változatokban az emberi populációban, elég ahhoz, hogy haploinsufficienciára utaljon. A SEC63 kevésbé korlátozottnak tűnik, bár ez azt is jelentheti, hogy csak recesszív módon hat.
A fentiek közül egyik sem ad választ arra a kérdésre, hogy miért léteznek GPI-horgonyzott fehérjék. A régi sejtbiológia órámról egyébként kimaradt egy részlet: valójában létezik a membránfehérjéknek egy hatodik osztálya, az úgynevezett tail-horgonyzott (TA) fehérjék , amelyeknek csak egy hidrofób C terminusuk van, amely a membránba tapad, de a másik oldalon nem áll ki. Miért ne lehetne az összes GPI-horgonyzott fehérje egyszerűen TA-fehérje? Miért fejlesztettek ki a sejtek egy ilyen bonyolult útvonalat, hogy ehelyett egy cukor-zsír horgonyt szintetizáljanak, és miért fejlesztették ki ilyen korán – a GPI horgonyok minden eukariótában jelen vannak, beleértve számos egysejtű kórokozót, amely az embert fertőzi.
A legtöbb áttekintés nem töltött sok időt ezzel a kérdéssel, valószínűleg azért, mert ez a legnehezebben megválaszolható kérdés. Maguk a GPI-horgonyzott fehérjék, amennyiben a natív funkcióik ismertek, a funkciók hatalmas skáláját képviselik – vannak enzimek (mint az AChE), sejtadhéziós molekulák (mint az NCAM1), az immunrendszerben a komplementet szabályozó fehérjék (CD59) és így tovább . Úgy tűnik, hogy legalább egy GPI-horgonyzott fehérje részt vesz a perifériás idegekben a mielin fenntartásában . De pontosan mit tudnak a GPI-horgonyzott fehérjék, amit más fehérjék nem? Egy áttekintés idéz néhány javasolt ötletet. Az egyik az, hogy a GPI-horgonyzott fehérjék jól képesek az átmeneti dimerizációra . Néhány tanulmány vizsgálta azt az elképzelést, hogy a homodimerizáció szerepet játszik a prionbiológiában , bár az ott használt modellrendszerek relevanciája az in vivo helyzetre még nem világos. Egy másik elképzelés szerint, mivel a GPI-horgonyzott fehérjék leválhatnak a sejtfelszínről, például az angiotenzin-konvertáló enzim (ACE) által, lokalizációjuk valamilyen dinamikus módon szabályozható. Itt is tudjuk, hogy a PrP le tud válni, nyilvánvalóan az ADAM10 enzim által, bár a PrP natív funkciójában betöltött szerepe még nem világos. Egy harmadik elképzelés, amelyről talán a legtöbbet hallottam, hogy a GPI-horgonyzott fehérjék szelektíven “lipidtutajokban” gyűlnek össze. Talán ez a legcsábítóbb magyarázat, mert mindenféle tovagyűrűző hatást el lehet képzelni, ahol e fehérjék megnövekedett hatékony helyi koncentrációja több kölcsönhatást tesz lehetővé, és így tovább. Az egyik áttekintés azonban rámutatott, hogy egy figyelmeztetés, hogy a lipidtutajok még mindig inkább absztrakt elképzelés, mint konkrét dolog – bár funkcionálisan a detergensek oldhatatlansága határozza meg őket, és a legtöbb ember úgy írja le őket, hogy szfingomielinben és koleszterinben gazdagok, nincs általánosan elfogadott definíció arra, hogy mi a lipidtutaj és mi nem az, és az empirikus bizonyítékok arra utalnak, hogy ezek sokkal kisebbek és átmenetibbek lehetnek, mint a legtöbb ember gondolja.
Ezzel az olvasmánnyal a kezemben elindultam, hogy szerezzek egy listát ezekről a fehérjékről, és elvégezzek rajtuk néhány elemzést, hátha jobb képet kapok arról, hogy milyenek.
analízis
Az Uniprot 173 humán GPI-horgonyzott fehérjéről tartalmaz egy listát. Ezek 140 génszimbólumot képeztek le, ami 135-re csökkent, miután lefuttattam ezt a szkriptet, hogy frissítsem a jelenleg HGNC által jóváhagyott fehérjekódoló génszimbólumokra. A 135 génszimbólumot tartalmazó végleges lista itt található.
Az Uniprot nem ad semmilyen információt arról, hogy az annotációkat hogyan generálták, bár jelentős mértékű manuális kurációnak kell lennie. Összehasonlításképpen, Andrew kiásott egy sor csinos cikket is, amelyek a PI-PLD-t vagy a PI-PLC-t, két GPI-horgonyokat hasító enzimet használták a GPI-horgonyzott fehérjék empirikus izolálására a sejtekből . Az ezekből a papírokból származó listák kombinálásával és a jelenlegi génszimbólumokhoz való hozzárendeléssel 107 gént kaptunk. Ezek közül néhányat véletlenszerűen szúrópróbaszerűen ellenőriztünk. Ezek között voltak olyan jól ismert GPI-horgonyzott fehérjék, mint a glipikán-1 (GPC1) és a neurális sejtadhéziós molekula (NCAM1), amelyek mindegyike a jelentések szerint kölcsönhatásban áll a PrP-vel . De számos olyan gén is jelen volt, amelyekről az irodalomban nem tűnt ismertnek a GPI-horgonyzás, mint például a VDAC3, amelyek közül néhány egyszerűen csak nagyon gyakori fehérje vagy más okokból fals pozitív eredmény. Eközben vannak nyilvánvaló hamis negatív források is: olyan gének, amelyek egyszerűen nem fejeződtek ki a vizsgált sejtvonalban, vagy nem voltak elég bőségesek ahhoz, hogy tömegspektrométerrel kimutathatók legyenek, és a PrP paralógjai, az SPRN és a PRND nem szerepeltek a listákon. Összességében 51 gén szerepelt mindkét listán, ami rendkívül szignifikáns gazdagodás (OR = 217, P < 1 × 10-84), ami segít megnyugtatni engem, hogy az Uniprot annotációi összhangban vannak az empirikus adatokkal. A további elemzésekhez azonban úgy döntöttünk, hogy az Uniprot-listát választjuk, mivel az érzékenyebbnek és specifikusabbnak tűnik.
Ezzel a listával felvértezve szerettem volna megnézni, hogyan állnak a GPI-horgonyzott fehérjék. A PrP egy egy exonos, rövid (érett formájában 208 aminosav), nem esszenciális, széles körben expresszálódó fehérje. Ezek a jellemzők tipikusak vagy atipikusak egy GPI-horgonyzott fehérje esetében?
Kiderült, hogy a GPI-horgonyzott fehérjék a térképen mindenhol megtalálhatók, ugyanolyan változatosak minden általam vizsgált dimenzióban, mint bármely más fehérjecsoport.
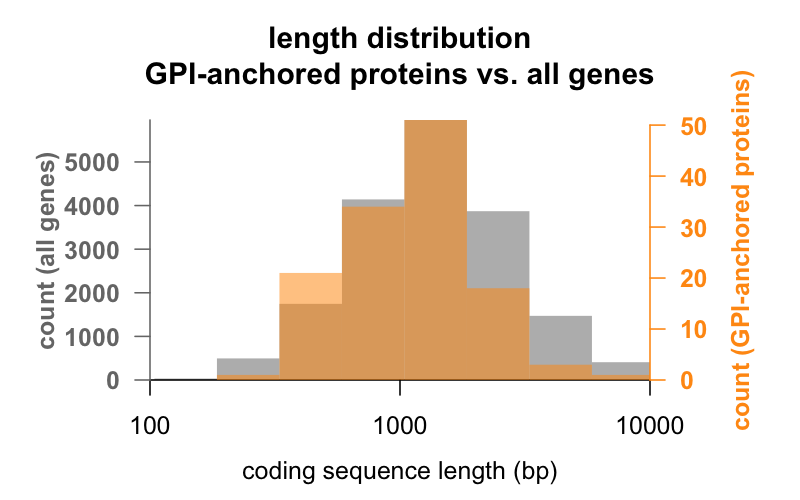
Először is, a hossz. Az alábbiakban az összes gén és a GPI-horgonyzott fehérjéket kódoló gének kódoló szekvencia hosszának bázispárokban kifejezett hisztogramjai vannak egymásra helyezve. A GPI-horgonyzott eloszlás épphogy balra tolódik. A GPI-horgonyzott fehérjék génjének átlagos kódoló szekvenciája 1301 bp, míg az átlagos géné 1729 bp, de ez az átlagkülönbség kicsi az egyes csoportokon belüli eltérésekhez képest. A PrP a mindössze 762 bp kódoló szekvenciájával határozottan a kicsi oldalon áll, bár semmiképpen sem számít kiugrónak egyik csoportban sem – a CD52, mindössze 186 bázispárnyi szekvenciájával és érett formájában nyilvánvalóan csak 12 aminosavval , a legkisebb GPI-horgonyzott fehérje.
Mi a helyzet az exonok számával? A GPI-horgonyzott fehérjéknek valóban átlagosan valamivel kevesebb exonjuk van, mint az összes génnek (átlagosan 7,8 vs. 10,1), ami összhangban van a fent említett enyhe hosszeloszlási különbséggel, de a legtöbbjük több exonból áll. A PrP itt is a kicsi oldalon áll: mindössze hat olyan GPI-horgonyzott fehérje van, amelynek csak 1 kódoló exonja van, és ezek közül három a PrP és két paralógja, a Sho és a Dpl. (A másik három gén a GAS1, a SPACA4 és a mesés nevű OMG).
A következőkben a funkcióvesztéses korlátozást vizsgáltam. A korlátozás annak a mértéke, hogy egy gén milyen erős természetes szelekciónak van kitéve, az alapján, hogy az általános populációban a mutációs rátákon alapuló várakozásokhoz képest mennyire fogyatkozik meg mondjuk a nonszensz, a frameshift és a splice site variáció. Ez a mérőszám rövid gének esetében nem nagyon értelmezhető, mind statisztikai okokból (a várható mutációk száma rövid gének esetében alacsony, így nehéz számszerűsíteni a kimerülést), mind biológiai okokból (az egy exonból álló gének nincsenek kitéve nonsense-mediált bomlásnak, így nehezebb eldönteni, hogy a fehérje-trinkelő változatok valóban “funkcióvesztésnek” minősülnek-e vagy sem). De mivel a legtöbb GPI-horgonyzott fehérje nem olyan rövid, mint a PrP, úgy gondoltam, érdemes megnézni. Az eredmény: átlagosan a GPI-horgonyzott fehérjék csak valamivel kevésbé korlátozottak, vagyis több a funkcióvesztéses variáció várható mennyisége, mint az átlagos géneknél. Az átlagos gén a funkcióvesztéses variáció 47%-ával rendelkezik, míg a GPI-horgonyzott fehérjék 56%-ával. De mint mindenben, itt is széles a szórás mindkét táborban. A GPI-horgonyzott fehérjék esetében az egyik végén van az abszolút korlátozott ACHE (17 LoF várható, de egyet sem figyeltek meg), a másik végén pedig számos olyan gén, amely egyáltalán nem szelektálódik a funkcióvesztés ellen – a CNTN6, CD109, TREH és MSLN csak néhány példa. A PRNP az utóbbi táborba tartozik, ha kizárjuk az ≥145-ös maradékokat, ahol a fehérjecsonkoló változatok funkciógyarapodást okoznak.
Végezetül elgondolkodtam azon, hogy hol fejeződnek ki a GPI-horgonyzott fehérjék. A PRNP az agyban a legmagasabb, de mindenhol expresszálódik. Ez tipikus? Letöltöttem a teljes GTEx v7 “gene median tpm” összefoglaló fájlt (2016. január 15.), ahol minden sor egy gén és minden oszlop egy szövet, a cellák pedig RPKM-ek – RNA-seq olvasások kilobázisonként exononként egymillió leképezett olvasásonként. Az ezzel az adatkészlettel való munka némi finomkodást igényelt. Hallottam, hogy egyes bioinformatikusok a <1 RPKM-et “nem kifejezettnek” tekintik, de az expressziós mátrix ritka – a legtöbb gén a legtöbb szövetben nem kifejezett -, így az 1 RPKM alatti zaj dominálhat, ha csak a nyers RPKM-eket ábrázoljuk. Eközben a génexpresszióra logaritmikus skálán kell gondolni, mivel a gének egy szövetben <1 RPKM és >10 000 RPKM között változhatnak, így ha mindent lineáris skálán tekintünk, akkor a néhány igazán magasan expresszált gén/szövet kombináció is dominálhat, így a mátrix még ritkábbnak tűnik, mint amilyen valójában. Ezért a mátrix log10-ét vettem, és az eloszlást , így az általam használt lila skála 1 – 10 – 100 – 1.000 – 10.000 RPKM-nél csonkoltam meg. Ezután az Uniprot GPI-horgonyzott fehérjékre osztottam. Ennek vizualizálásához életemben először készítettem egy hőtérképet. Gyakran láttam már ilyeneket a papírokban, és általában nem szólnak hozzám, de itt a célom csak az volt, hogy érzékeljem az expressziós mintázatot, és miután egy kicsit játszottam, ez volt az, ami a legtöbb betekintést adta nekem. A hőtérkép elve az, hogy a sorok és oszlopok úgy vannak csoportosítva, hogy a hasonló dolgok együtt járnak. Így például az összes agyszöveti oszlop egymás után egy foltban sorakozik az x tengelyen, és az összes erősen agyi expressziójú gén egymás után egy foltban sorakozik az y tengelyen, így ezek metszéspontja egy sűrű lila téglalapot alkot, amely úgy értelmezhető, hogy “létezik egy olyan géncsoport, amely főként agyi expressziójú”.
Az érdeklődő olvasók megtekinthetik a hőtérkép teljes vektorgrafikus PDF formátumát, de hogy közvetlenebbé tegyük, íme egy kézzel jegyzetelt változat, amely az érdekes klasztereket nevezi meg:

A válasz tehát nem – a legtöbb GPI-horgonyzott fehérje nem rendelkezik olyan expressziós mintázattal, mint a PRNP. A PRNP egyike a maroknyi magasabban és szélesebb körben expresszálódónak, amely a CD59, a LY6E, a GPC1 és a BST2 mellett a hőtérkép teteje közelében szerepel. A legtöbb GPI-horgonyzott fehérje kifejeződése kisebb vagy erősebben szövetre korlátozódik, egyesek csaknem kizárólag az agyban fejeződnek ki, mások csaknem kizárólag az agyban nem, és más kisebb csoportosulások főként specifikus szövetekhez, például a herékhez tartoznak, mint például a PrP paralógja, a PRND, amelynek kiütése férfi sterilitást okoz.
következtetések
A GPI-horgonyzott fehérjék szinte bármilyen méretűek lehetnek, szinte bármilyen szövetben kifejeződhetnek, és nyilvánvalóan szinte bármilyen funkcióval rendelkezhetnek, amennyiben a funkcióik ismertek. Sok GPI-horgonyzott fehérje nagyon világos natív funkcióval rendelkezik, de ezek a funkciók sokfélék, és nem világos, hogy miért van szükségük GPI-horgonyzásra, különösen azért, mert sok ilyen fehérje létezik nem-GPI-horgonyzott izoformában is. Eközben más GPI-horgonyzott fehérjék, köztük a PrP esetében eleve elég keveset tudunk a natív funkcióról, így nehéz még csak találgatni is, hogy a natív funkcióhoz miért van szükség GPI-horgonyzásra. Az általam elvégzett elemzések vagy az általam olvasott áttekintések egyike sem tudott egységesítő elvet felállítani arra vonatkozóan, hogy miért létezik ez a lehorgonyzási mechanizmus, vagy hogy miért van szükségük ezeknek a fehérjéknek erre. Számos hipotézis létezik arra vonatkozóan, hogy a GPI-horgonyzott fehérjék miért egyedülállóak, beleértve a lipid tározókat, a homodimereket és a leválást. Mindegyik hipotézisnek lehet némi alapja. De a nap végén úgy tűnik, hogy a válasz nem egy heuréka pillanat, hanem, mint a biológiában oly sok minden, különböző dolgok prózai keveréke.
A kód és a nyers adatfájlok az ebben a bejegyzésben szereplő elemzésekhez itt találhatók.