Replikation
Kopiering: Cool for DNA, Cool for os
Da nogle af Shmoops klassekammerater gik i skole, tog deres lærere dem altid i at kopiere. Men klassekammeraterne fortalte deres lærere, at det lå i deres DNA at kopiere. Hun kunne ikke argumentere imod den logik, så de kom aldrig i problemer. Sikke en fin lille historie.
Hvad er det for en parallel, vi forsøger at drage med denne ikke-så-subtile og skingre fortælling? DNA kopierer hele tiden, og det skal det også, hvis det vil blive ved med at lave alle de celler. Og, åh, hvor det dog gør det! DNA-replikation er kopiering af DNA, så de replikerende celler har nok DNA til datterceller, eller de nye celler, der stammer fra den oprindelige celle. Celledeling, eller dannelsen af en celle til to nye celler, sker gennem mitose for ikke-kønsceller, eller de celler, der ikke er involveret i … sex, og meiose for kønsceller, eller de celler, der kommer ned og bliver beskidte hurtigere end Vinny og Pauly D på en god aften i Karma.
Det er bare en fase: Mitose eller meiose
Trods celler findes i alle former og størrelser, ligesom en Benetton-reklame, men der er grundlæggende to typer celler:
- Kønsceller (nej, ikke de der ting i Amsterdam)
- Somatiske celler (ikke-kønsceller, som heller ikke er de der ting i Amsterdam)
Kønsceller er de celler, der producerer afkom hos eukaryoter, der formerer sig seksuelt, og de er opdelt i følgende:
Kønsceller er de celler, der producerer afkom hos eukaryoter, der formerer sig seksuelt, og de er opdelt i følgende:
- Spermceller, der indeholder den mandlige halvdel af DNA
- Egceller, der indeholder den kvindelige halvdel af DNA

Der er fire hovedstadier i livscyklusen for de fleste eukaryote celler (undtagen kønsceller).
Disse kaldes
- Gap 1 (eller G1) fase
- Syntese (eller “S”) fase
- Gap 2 (G2) fase
- Mitose
Og nej, The Gap sponsorerer ikke mitose. G1-, S- og G2-fasen kaldes normalt for “interfase”, mest af folk, der studerer mitose. Selvom, hvis man får dem alene, kalder de det “kedelig fase.”
G1
G1 er den fase i den eukaryote livscyklus efter mitose og før DNA-syntese, som er S-fasen. I løbet af mitosen er det meste af den aktivitet, der ikke er mediose, blevet lukket ned, og G1 er den periode, hvor alt starter forfra. G1 er som en sindssyg søvngænger, der vækker cellen ved at sige: “Hej, jeg har skåret kuponer ud hele natten, det er på tide at vågne op og lege med mig!”
S-fase
S-fasen er livscyklussens DNA-replikationsfase. Vi vil tale mere om dette senere, men hvis du ikke kan vente, så klik videre. I denne fase fordobler cellen mængden af DNA for at lave nok DNA til dattercellerne, og cellen går fra 2n til 4n, hvor n er antallet af sæt kromosomer, eller ploidy, og cellen går fra 2n til 4n. De fleste somatiske celler er 2n eller diploide, mens kønsceller er haploide, hvilket betyder, at de har halvt så meget DNA som somatiske celler (almindelige celler).
G2-fase
G2 er cellecyklusens Jan Brady. Alle snakker altid om “Mitose, Mitose, Mitose!”, og ingen bekymrer sig om stakkels G2. Det er den periode i cellecyklussen, hvor der sker mere proteinsyntese, og hvor der laves mikrotubuli, som er vigtige for mitose. “Det er det hele?” Spørger du? Jep. Ikke så vigtigt, og det er derfor, at mange celler springer dette trin over. Frøceller springer det over, og det samme gælder for mange kræftceller. Stakkels G2…
Mitosis
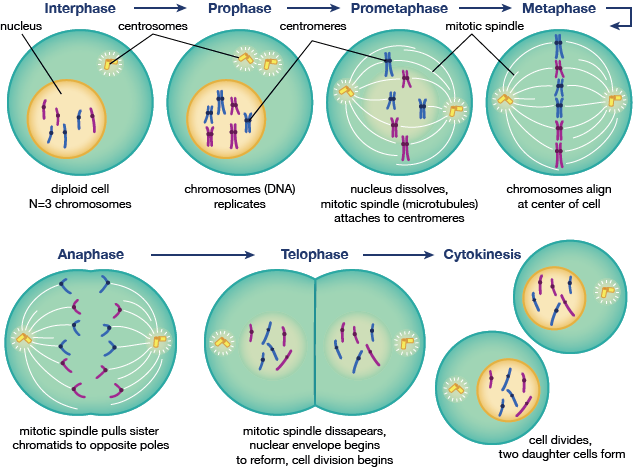
Mitosis er det trin i cellecyklus, der er så vigtigt, at det har sin egen cyklus. Mitose begynder efter G2 og starter med profasen, eller kondensering af kromatin til kromosomer og centrosomer, der bevæger sig mod modsatte poler i cellen. Dette trin efterfølges af prometafasefasen, et mellemstadium mellem profasen og metafasen, hvor kernehulen nedbrydes, og kromosomerne begynder at samle sig på mikrotråde, der forbinder centrosomerne. Metafase er det tidspunkt, hvor alle kromosomerne er rettet op i midten, hvilket efterfølges af anafase, hvor kromosomerne deles i to, og datterkromatider bevæger sig til den nærmeste centrosompol. Telofase og cytokinese er de sidste trin i celledelingen og genforeningen af kernen i henholdsvis to datterceller.
Meoisis

Meiose er M-fasen for kønsceller. Den begynder som mitose, og efter produktionen af de to første datterceller fortsætter den en anden mitosecyklus. De 2n datterceller deler sig yderligere og producerer 4 datterceller, og en masse overpris bryllup, hver med n kromosomer.
I begyndelsen…DNA-replikationens oprindelse
Hvis DNA aldrig replikerede sig, ville meiose og mitose langsomt halvere genomets størrelse, indtil hver celle ville dø, hvilket sandsynligvis ikke ville tage lang tid. Derfor er det vigtigt, at DNA fordobler sig selv for at tage højde for, at cellerne deler sig under mitose/meiose. DNA-replikation svarer til RNA-transkription.
Nedenfor er en tabel, der sammenligner de to:
| DNA-replikation | RNA-transskription | ||
| Produkt | Dobbelt-strenget DNA | mRNA | |
| Enzym | DNA-polymerase | RNA-polymerase | |
| Nucleotider Tilføjet | Deoxynukleotidtrifosfater | Nukleotidtrifosfater | |
| Template Strand | Både | Både | Både, men transskriberer kun fra antisense-strengen |
Hvad kalder du de steder, hvor replikationen starter? Oprindelser. Vi ville ønske, at de havde kaldt det Star Wars Episode I: The Phantom Menace, men det navn var allerede taget.
Originerne er forskellige alt efter typen af DNA. De fleste eukaryote celler og nogle bakterier og vira har lineært DNA, hvilket betyder, at der er en begyndelse og en slutning på DNA-sekvensen. Derfor er replikationsoprindelsen i 5′-enden af det lineære DNA. De fleste bakterier og nogle vira har cirkulært DNA, hvilket betyder, at der er en specifik sekvens, som DNA-replikationsproteiner binder sig til (hvilket er noget analogt med RNA-transkription), og replikationen begynder fra dette sted.

DNA-replikation starter med en initiatorproteinbinding, som f.eks. dnaA i E. coli-bakterier, eller et kompleks af proteiner, som f.eks. origin recognition complex i gær. Initiatorernes funktion er at bryde de to DNA-strenge fra hinanden, således at replikationen kan begynde. Denne situation er ikke ulig en donkraft, der løfter din bil, når du skifter et dæk. (For det ved vi alle sammen, hvordan man gør, ikke?) Hvis du ikke ved, hvordan man skifter et dæk, så ring til et kranvognsselskab og spørg dem, hvordan de gør det. Når de fortæller dig hvordan, skal du skrige: “
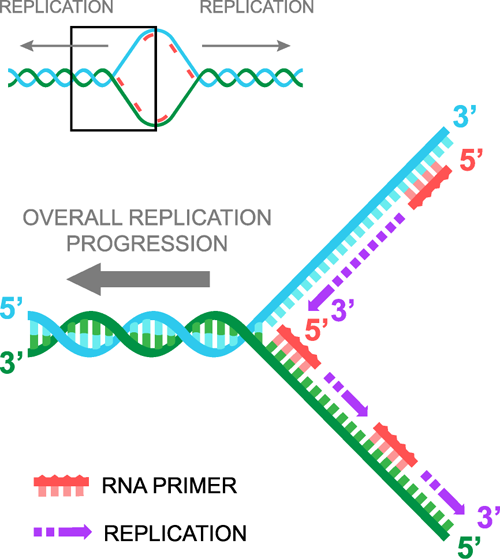
Når DNA’et er blevet skilt ad, kommer RNA-præmier ind, en binder sig til hver DNA-streng, og så fortsætter DNA-replikationen i en 5′-til-3′-retning. Det er vigtigt at indse, at der sker to 5′ til 3′-replikationer samtidig. Disse to replikationer kaldes hver især en replikationsgaffel. Ikke at forveksle med duplikationsskeen.

Passér replikationsgaflen
Når man deler DNA, har vi en streng, der står 5′ til 3′, når vi ser fra venstre til højre, som vi vil kalde den “bagudrettede” streng, og en komplementær streng, der står 3′ til 5′, når vi ser fra venstre til højre, som vi vil kalde den “førende” streng. Når vi tilføjer RNA-primere, binder den ene primer sig derfor til den ledende streng, og den anden primer binder sig til den bagudrettede streng. Den primer, der binder til den ledende streng, er identisk med sekvensen af den bagvedliggende streng, mens den primer, der binder til den bagvedliggende streng, er identisk med sekvensen af den ledende streng. Vi håber ikke, at dit hoved er eksploderet endnu.
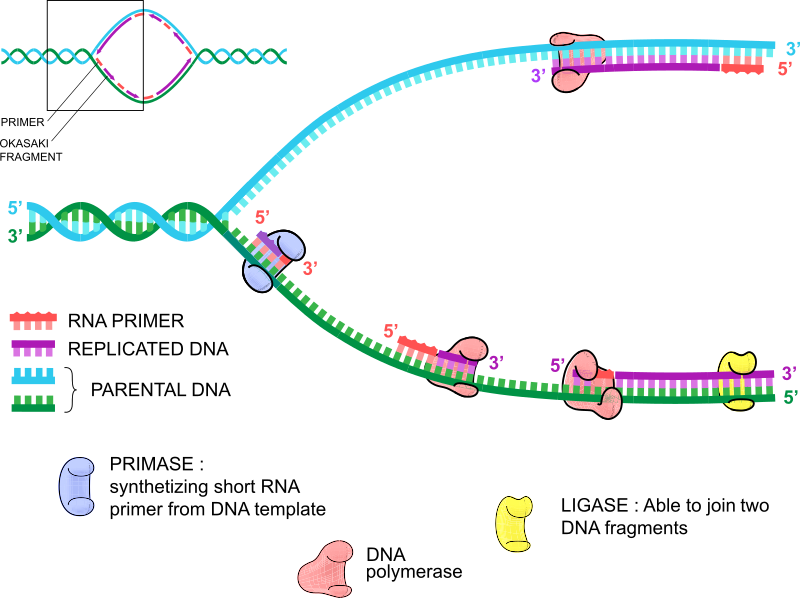
Tænk på DNA-replikation som din venstre og højre hånd. De er i bund og grund de samme, men har subtile forskelle. Din venstre hånd har tommelfingeren som den femte finger, hvis du tæller fra venstre til højre, mens din højre hånd har tommelfingeren som den første finger. Se? Undskyld, hvis vi har gjort dig forvirret. Derfor replikerer begge DNA-strenge sig ligesom dine hænder på samme måde, men der er en subtil forskel. DNA-replikation fungerer kun i 5′ til 3′-retningen, så to DNA-polymerase-molekyler binder RNA-primere og begynder at replikere i modsat retning. Replikationen af den forreste streng driver replikationsprocessen, fordi RNA-primeren driver replikationen af 5′- til 3′-strengen.
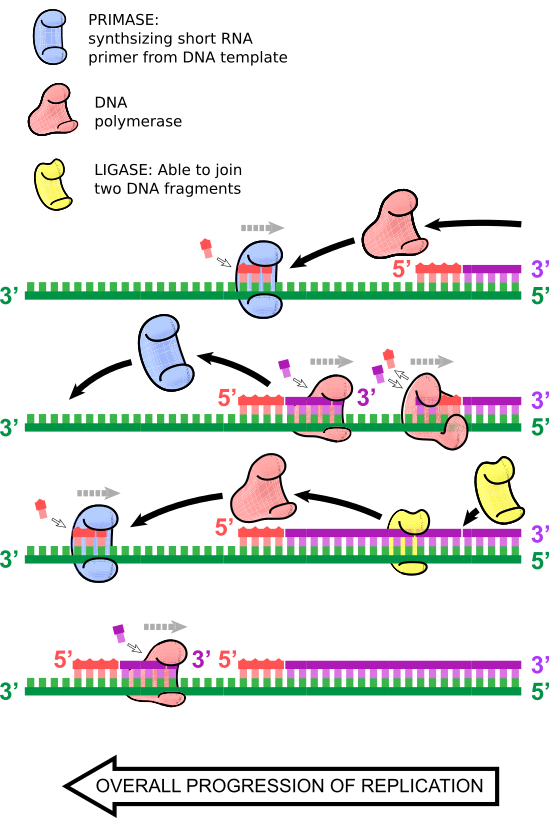
“Men Shmoop, hvordan replikerer man den ‘bagvedliggende’ streng?”, vil du måske spørge.
Som navnet antyder, fortsætter DNA-replikationen af den ledende streng i et stykke tid, indtil en ny RNA-primer indsættes, og en anden DNA-polymerase primerer fra denne anden laggende RNA-primer og replikerer DNA, indtil den rammer den første RNA-primer eller den tidligere strækning af dobbeltstrenget DNA. Her falder DNA-polymerasen fra, og DNA-ligase samler disse DNA-fragmenter. Denne proces fortsætter samtidig med syntesen af den ledende streng, så der er mange små replikerende dobbeltstrengede DNA-fragmenter på den forsinkede streng. Disse kaldes Okazaki-fragmenter, efter ægteparret Reiji og Tsuneko Okazaki, som opdagede dem.
Er vi færdige endnu?
DNA-replikation slutter i bakterier og vira med cirkulære genomer, når de to replikationsgafler møder hinanden på den modsatte side af kromosomet fra replikationsoprindelsen i en storslået kyllingespil. Det modige replikationsterminatorprotein træder ind og sætter DNA-replikationen på pause, så de ikke støder ind i hinanden. Termineringsmekanismen fører imidlertid til, at to cirkulære kromosomer sidder fast til hinanden som tryllekunstnerringe. Topoisomeraseenzymer, såsom DNA-gyrase og topoisomerase IV, som er ansvarlige for at knække DNA, skærer midlertidigt det ene kromosom over, så det andet kromosom kan passere igennem. Dermed ødelægger vi trylletricks for alle.
Animation af lineær kromosomforkortning
Lineære kromosomer har et andet sæt problemer, for med disse fyre når vi aldrig frem til en fuldstændig replikation af det fulde kromosom. Hvert replikationstrin forkorter DNA’et en lille smule mere. Bakterier og vira med lineære kromosomer omgår dette problem ved enten
- at starte replikationen i midten af kromosomet, eller ved
- at primere replikationen med et protein i stedet for en RNA-primersekvens.
Eukaryoter er imidlertid ikke i stand til at gøre nogen af disse to ting, og de har udviklet terminatorsekvenser kaldet telomerer, der omgår problemet med DNA-forkortning.

Enderne af lineært eukaryote DNA kaldes telomerer, og selv om de regelmæssigt forkortes ved hver replikationscyklus, løser enzymer kaldet telomeraser problemet med DNA-forkortning. Telomerer er meget repetitive, og telomeraser tilføjer korte sekvenser for at forlænge enden af DNA’et, så de ikke bliver for korte, og så vigtige dele af DNA-sekvensen ikke går tabt. Telomeraser er omvendte transkriptaser eller enzymer (man kan se det på -ase), der omdanner RNA til DNA. Se afsnittet “Spiderman og andre eksempler på rekombinant DNA” for at få mere at vide.
Telomeraser tager en kort RNA-sekvens, omdanner den til en dobbeltstrenget DNA-sekvens og sætter den fast til enden af DNA’et. Der er en minimumslængde, som telomerer skal opretholde, og hvis DNA-sekvensen er kortere end denne minimumslængde, kaldet Hayflick-grænsen, dør cellen. Telomeraser sikrer, at cellen kan fortsætte med at replikere og dele sig i det uendelige. Og overaktivitet af telomeraser er almindeligvis forbundet med kræft, da cellerne bliver udødelige, ligesom The Highlander.
Whoops, der blev begået en fejl: Du tænker sikkert: “DNA-polymerase synes, at den er så fantastisk, fordi den replikerer alt DNA i min krop, så jeg kan overleve og fortsætte med at gennemgå mitose og meiose.” OK, Vi er sikre på, at de fleste af jer ikke tænker det, men lad os spille med på den håndfuld mennesker, der tænker det.
Den DNA-polymerase er faktisk ikke så fantastisk! Den begår fejl ligesom du eller Shmoop – selv om vores fejl for det meste er produkter af for mange 5-timers energidrikke blandet med dybstegte Twinkies. DNA-polymerasen skal dog rette disse fejl, mens vi på Shmoop går i sukkerinduceret koma.

I hastværket ved DNA-replikationen tilføjes der sommetider en forkert base, der ikke baserparrerer korrekt med skabelonstrengen, til sekvensen. Derfor får vi underlige G-T- eller A-G-basepar. Vi husker alle, at normale par er G-C og A-T (Eh? Hvad er det?). Disse ukorrekte par er ikke gode for os, fordi det betyder, at vores gener muteres, og vi bliver mutanter. De færreste mutationer fører nogensinde til fantastiske ting som teleportation eller lasersyn … eller noget fra X-Men. Shucks.

Store eukaryoter som os, der replikerer langsomt og lever længe, ønsker ikke, at der skal ske mange mutationer. Hvorfor? Enhver mutation, der påvirker vores vækst, er normalt … ikke god … og vil skade os. Trist ansigt.
Men mindre organismer som bakterier, vira og endda fluer har korte livscyklusser i størrelsesordenen dage og nogle gange timer. De replikerer sig som gale, hvor hver ny generation vil have 100-1000-vis flere individer end den foregående generation. Derfor kan mutationer i deres tilfælde være mere fordelagtige. Nogle gange vil de gerne undslippe pesticider, antibiotika eller lægemidler, der er beregnet til at behandle/dræbe dem, hvilket er grunden til, at det er så svært at fremstille et lægemiddel, der kan forhindre os i at blive syge. Bakterier og vira bliver ved med at mutere og replikere. Og som vi allerede har bemærket, er der meget lidt fejlkontrol i deres polymeraser.
Hjernesnack
Her er en fin video om, hvordan DNA-replikation faktisk fungerer.
Kan du se DNA-polymeraserne? Det er de flerfarvede kugler, en til den forreste streng og en til den bageste streng. Det er de bagvedliggende polymeraser, der kommer ind og falder af, når dobbeltstrengen er færdiggjort.